

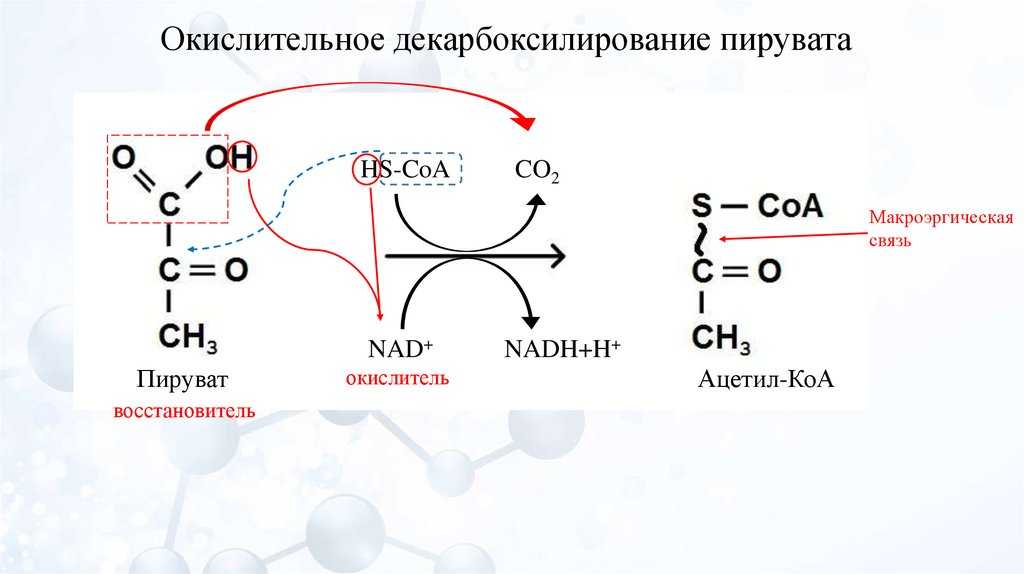
Пируватдегидрогеназный мульферментный комплекс
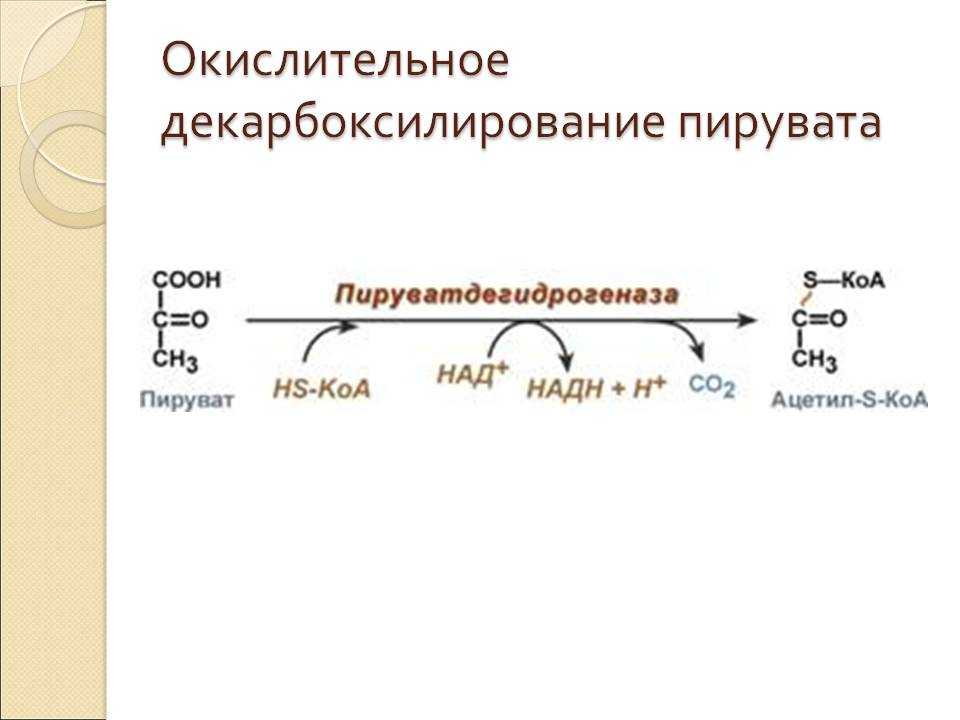
Суммарное уравнение отражает окислительное декарбоксилирование пирувата, восстановление НАД до НАДН и образование ацетил-SKoA.
Суммарное уравнение окисления пировиноградной кислоты
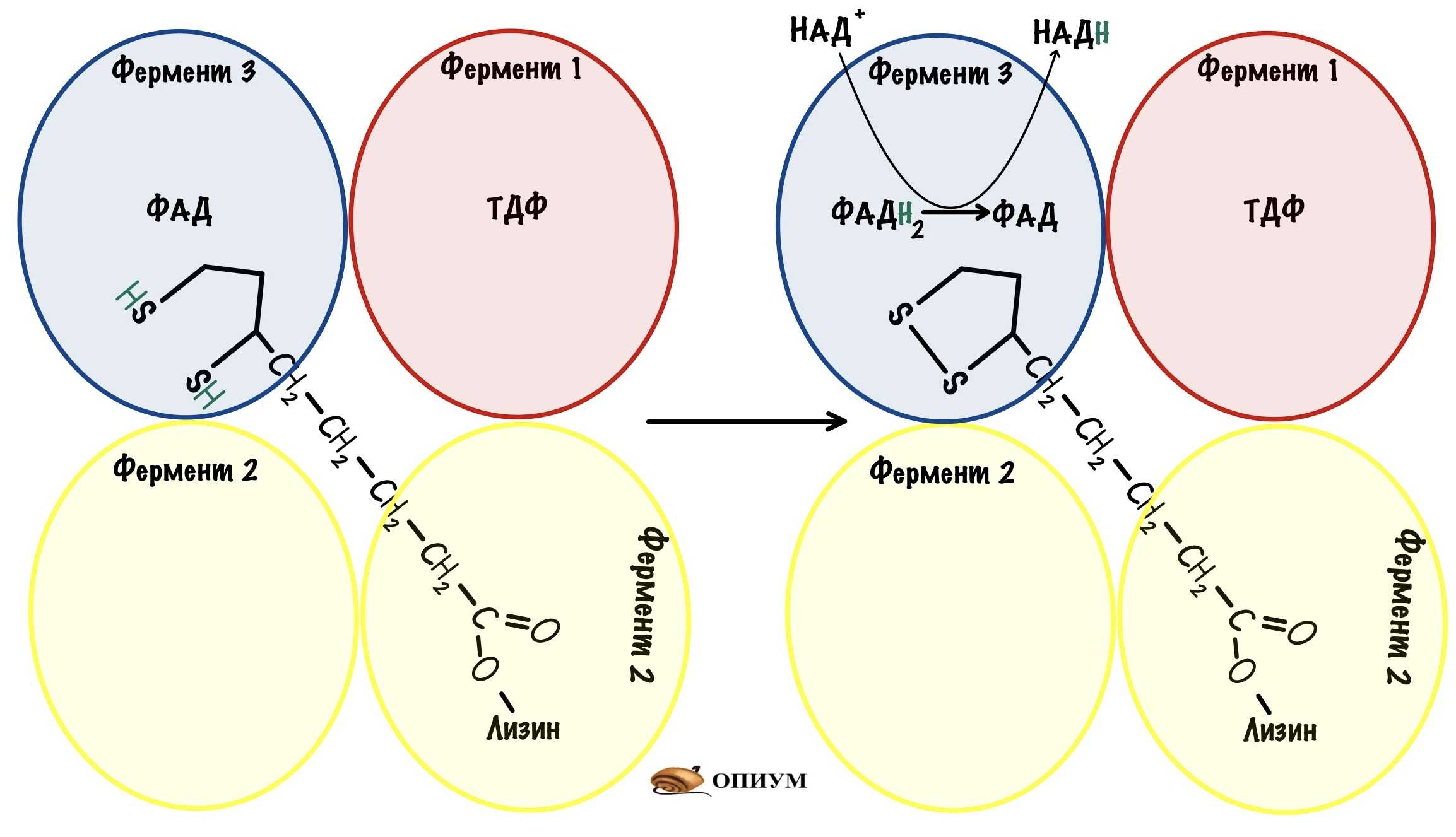
Превращение состоит из пяти последовательных реакций, осуществляется мультиферментным комплексом, прикрепленным к внутренней митохондриальной мембране со стороны матрикса. В составе комплекса насчитывают 3 фермента и 5 коферментов:
- Пируватдегидрогеназа (Е1, ПВК-дегидрогеназа), ее коферментом является тиаминдифосфат (ТДФ), катализирует 1-ю реакцию.
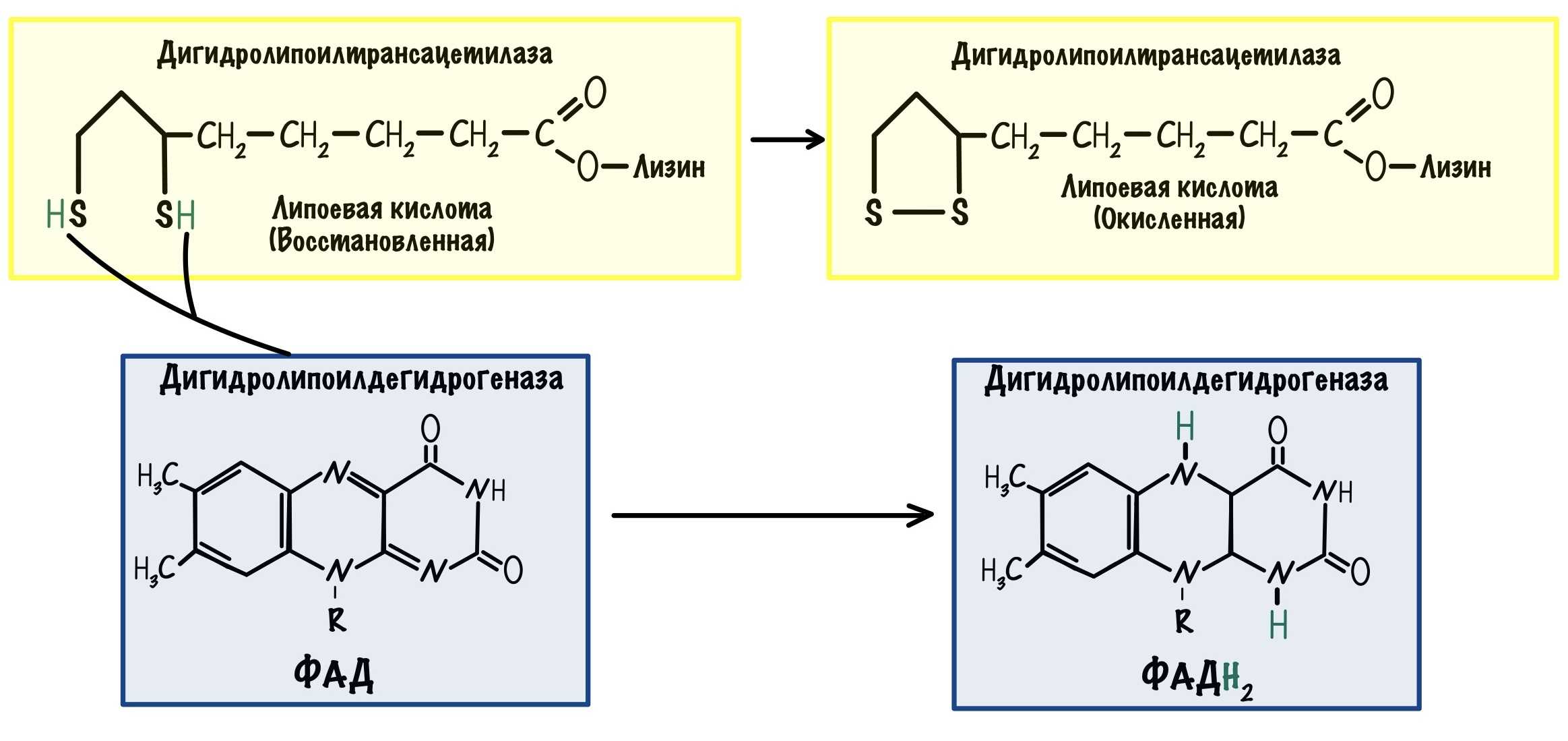
- Дигидролипоат-ацетилтрансфераза (Е2), ее коферментом является липоевая кислота, катализирует 2-ю и 3-ю реакции.
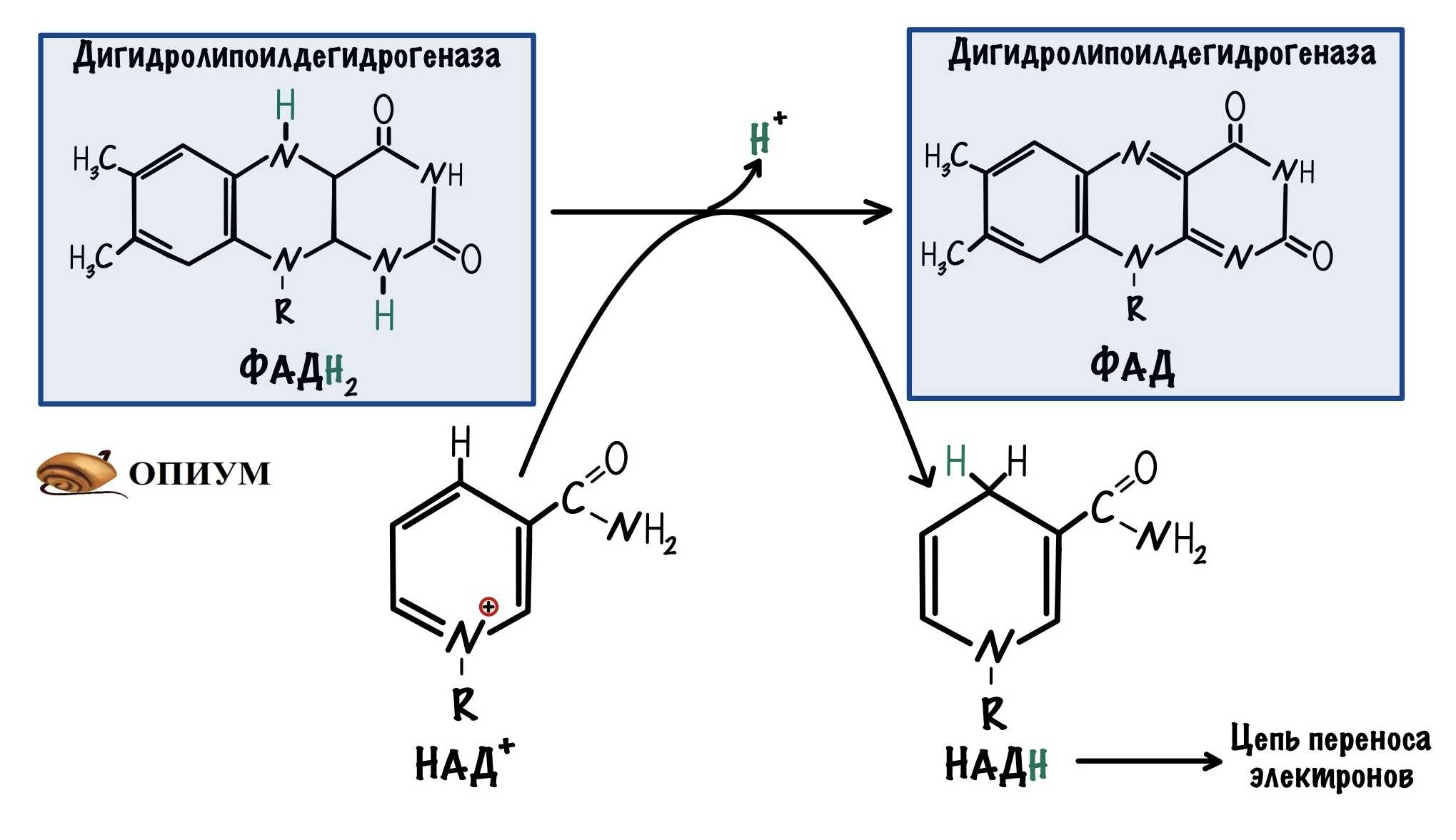
- Дигидролипоат-дегидрогеназа (Е3), кофермент – ФАД, катализирует 4-ю и 5-ю реакции.
Помимо указанных коферментов, которые прочно связаны с соответствующими ферментами, в работе комплекса принимают участие коэнзим А и НАД.
Суть первых трех реакций сводится к декарбоксилированию пирувата (катализируется пируватдегидрогеназой, Е1), окислению полученного гидроксиэтила до ацетила и переносу ацетила на коэнзим А (катализируется дигидролипоат-ацетилтрансферазой, Е2).
Реакции синтеза ацетил-SКоА
Оставшиеся 2 реакции необходимы для возвращения липоевой кислоты и ФАД в окисленное состояние (катализируются дигидролипоат-дегидрогеназой, Е3). При этом образуется НАДН.
Реакции образования НАДН
Регуляция пируватдегидрогеназного комплекса
Регулируемым ферментом ПВК-дегидрогеназного комплекса является первый фермент – пируватдегидрогеназа (Е1). Два вспомогательных фермента – киназа и фосфатаза обеспечивают регуляцию активности пируватдегидрогеназы путем ее фосфорилирования и дефосфорилирования.
Вспомогательный фермент киназа активируется при избытке конечного продукта биологического окисления АТФ и продуктов ПВК-дегидрогеназного комплекса – НАДН и ацетил-S-КоА . Активная киназа фосфорилирует пируватдегидрогеназу, инактивируя ее, в результате первая реакция процесса останавливается.
Фермент фосфатаза, активируясь ионами кальция или инсулином , отщепляет фосфат и активирует пируватдегидрогеназу.
Регуляция активности пируватдегидрогеназы
Таким образом, работа пируватдегидрогеназы подавляется при избытке в митохондрии (в клетке) ацетил-SКоА и НАДН, что позволяет снизить окисление пирувата и, следовательно, глюкозы в случае когда энергии достаточно.
Если АТФ мало или имеется влияние инсулина, то образуется ацетил-SКоА. Последний в зависимости от условий будет направляться либо в цикл трикарбоновых кислот с образованием энергии АТФ, либо на синтез холестерина и жирных кислот.
Анаплеротические реакции
Если мы забрали какие-то молекулы из ЦТК, то их концентрацию нужно восстановить. Иначе цикл не сможет работать. Реакции, с помощью которых восстанавливаются промежуточные соединения, называются анаплеротическими.
Можно восстановить любой субстрат цикла, потому что дальше он приведёт к образованию оксалоацетата. Это же цикл, правильно? Но таких реакций не слишком много. Выше мы видели, что альфа-кетоглутарат можно получить с помощью реакции трансаминирования. Есть и другие реакции, но на них останавливаться не будем.
Проще восстановить оксалоацетат, потому что в клетке много путей для его образования. В печени и корковом веществе почек оксалоацетат образуется с помощью пируваткарбоксилазы. Мы встречались с этой реакцией в глюконеогенезе.
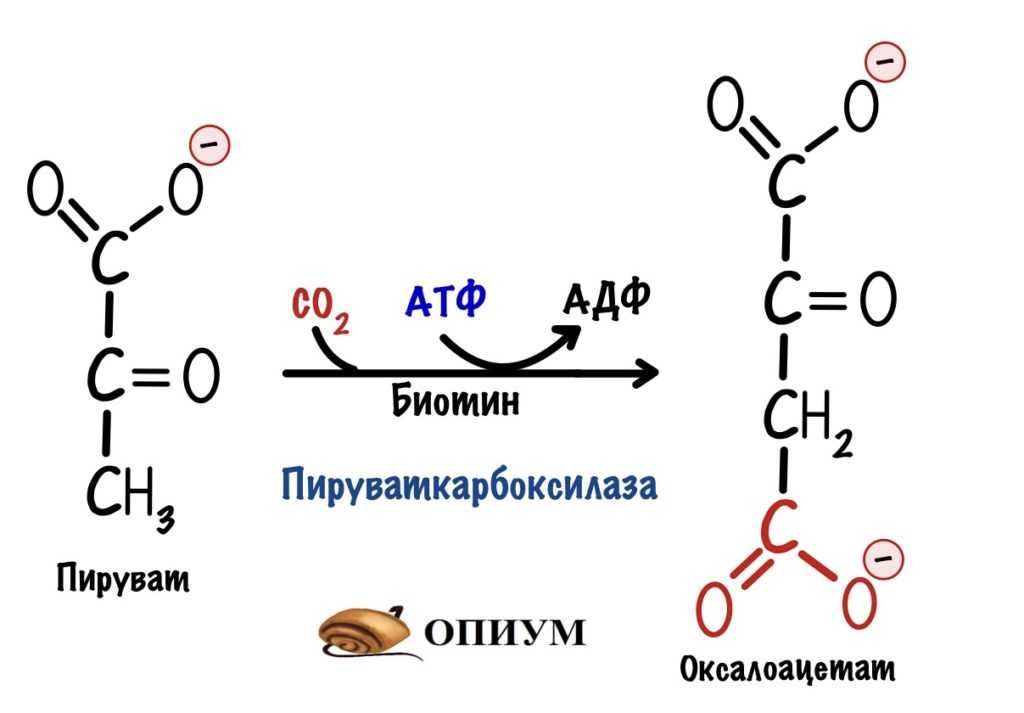 Восстановление концентрации оксалоацетата с помощью пируваткарбоксилазы. Эта реакция обратима, стрелочку не дорисовал.
Восстановление концентрации оксалоацетата с помощью пируваткарбоксилазы. Эта реакция обратима, стрелочку не дорисовал.
В сердце и скелетной мускулатуре оксалоацетат образуется из фосфоенолпирувата. Фермент — фосфоенолпируваткарбоксикиназа. В этой реакции разрушается макроэргическая связь фосфоенолпирувата, которая идёт на синтез ГТФ. Это реакция субстратного фосфорилирования. Но для нас важнее восстановить количество оксалоацетата, чем получить энергию.







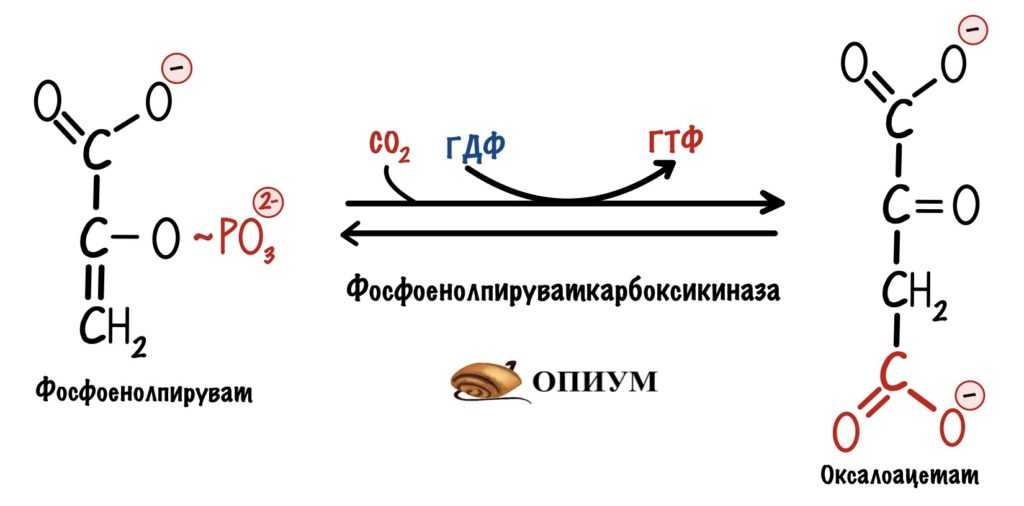 Эта реакция тоже была в глюконеогенезе, но шла в другую сторону.
Эта реакция тоже была в глюконеогенезе, но шла в другую сторону.
В большинстве тканей и органов есть малик-фермент. Он катализирует образование малата из пирувата. При этом его кофермент — НАДФH, а не НАДH. Я рассказывал про отличия между .
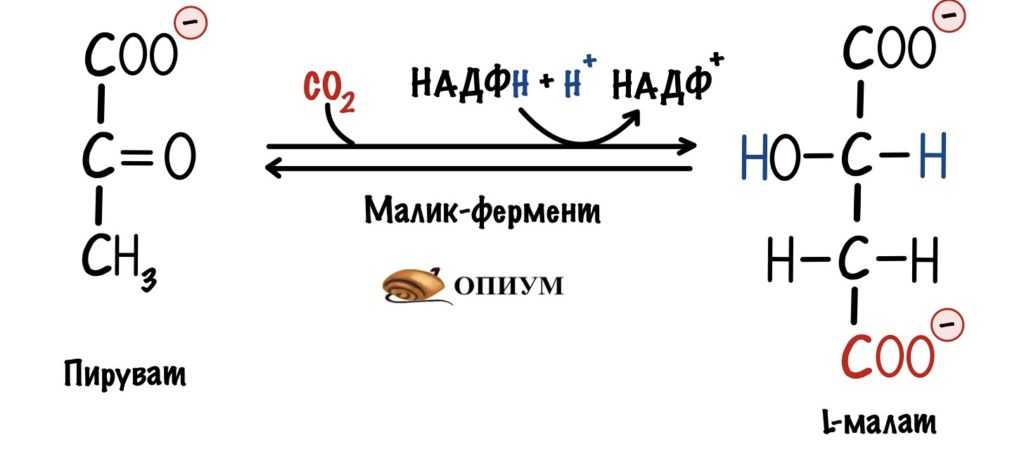 Малик-фермент
Малик-фермент
Малат от оксалоацетата отделяет всего одна реакция, которую катализирует малатдегидрогеназа. Мы уже говорили о ней выше.
Анаболическая роль ЦТК
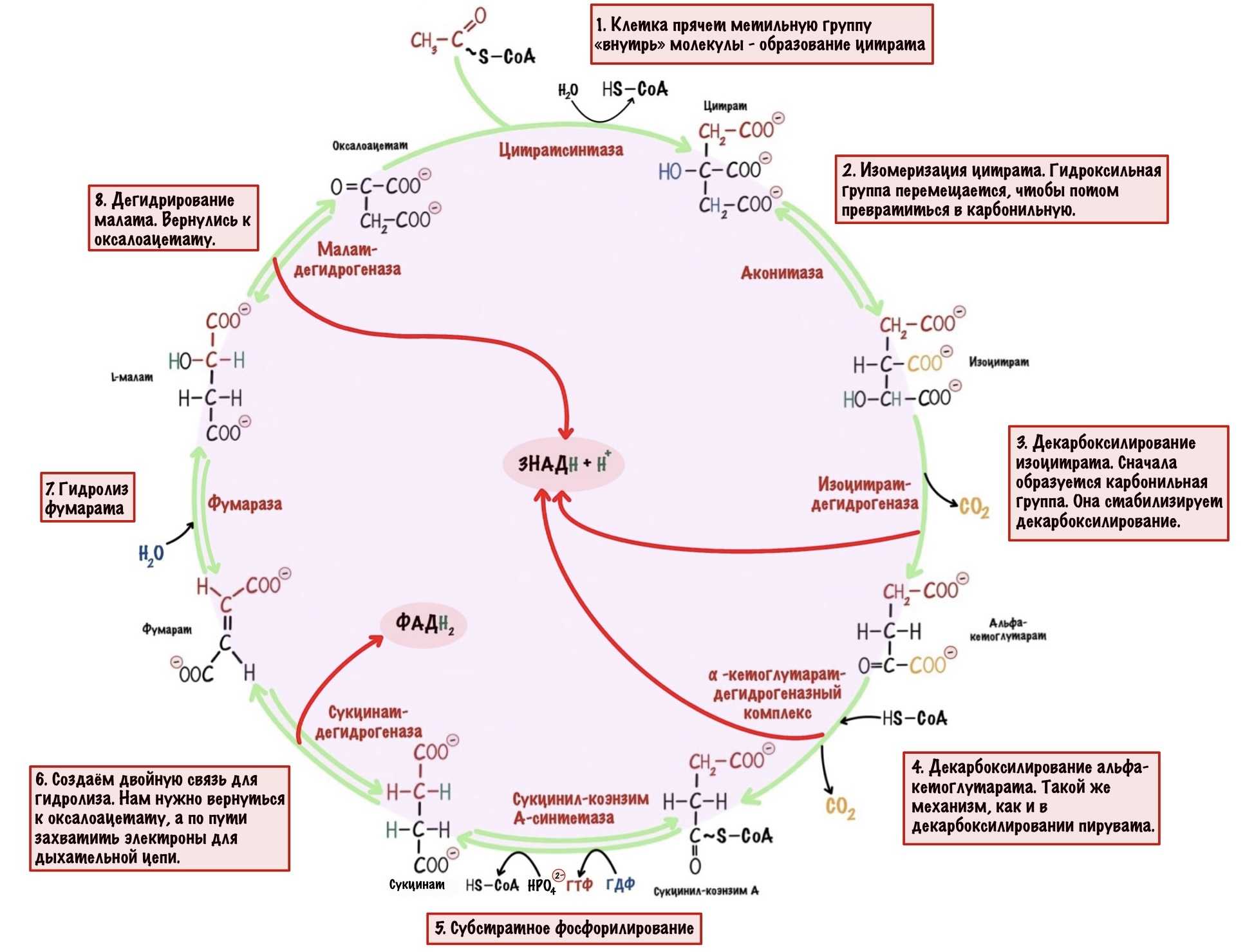
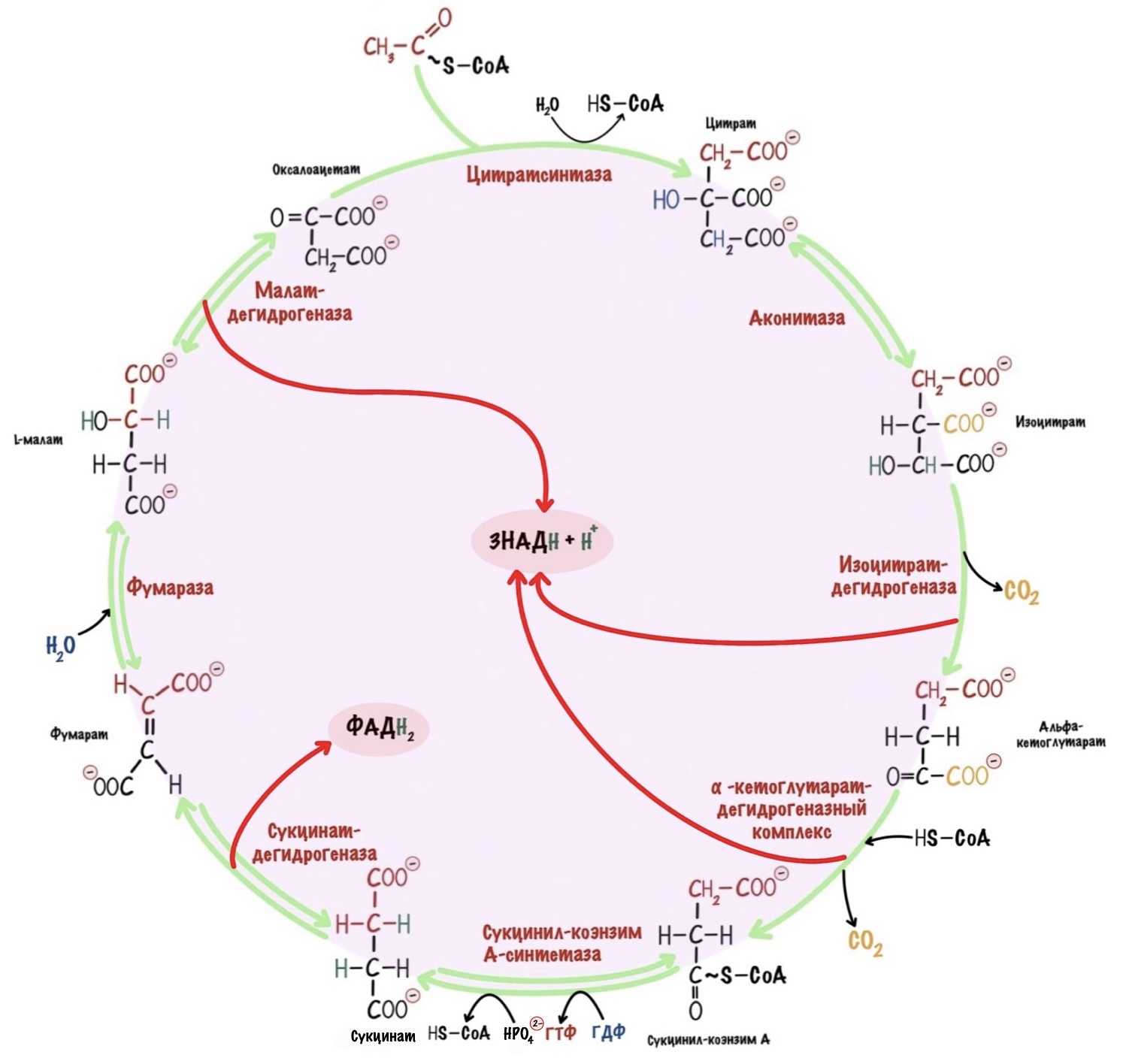
В самом начале статьи был разговор о том, что у ЦТК есть другая роль. А вот и она — анаболическая. В ЦТК восемь реакций. Они нужны не только для получения электронов. Часть молекул из цитратного цикла используются для анаболических процессов. Так что ЦТК — это амфиболический процесс, он сочетает катаболизм и анаболизм.
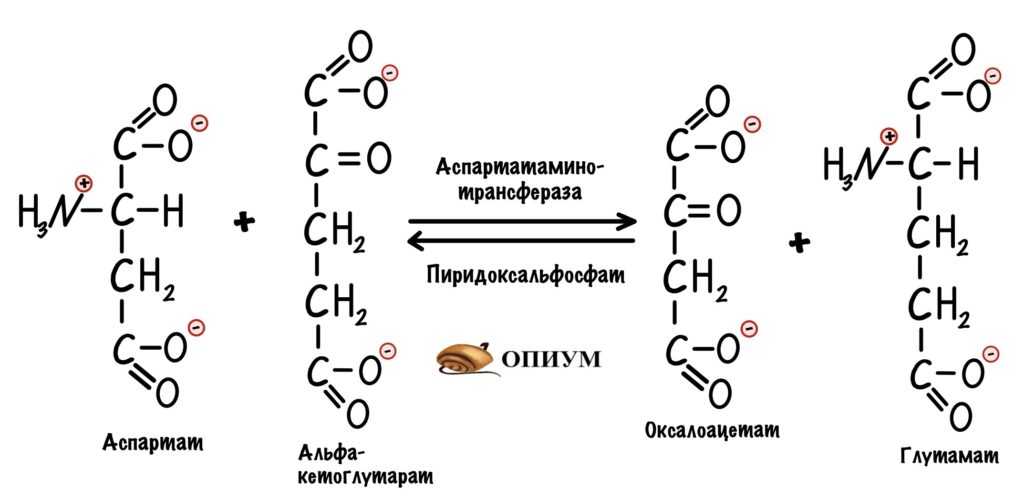
Сейчас разберём пару примеров, в которых цитратный цикл играет анаболическую роль. Начнём с первой реакции — образования цитрата. Нас интересует оксалоацетат, который вступает в неё. Из оксалоацетата можно получить не только цитрат. С помощью реакции трансаминирования оксалоацетата образуется аспартат.
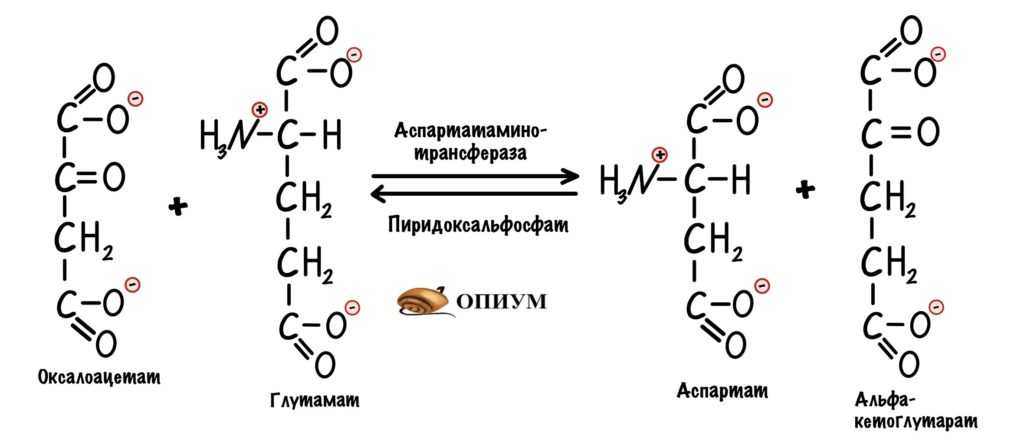 Использование оксалоацетата из ЦТК для анаболических процессов.
Использование оксалоацетата из ЦТК для анаболических процессов.
Аспартат используется для синтеза:
- Пиримидинов;
- Белков;
- Мочевины;
- Аспарагина
Если вспомнить глюконеогенез, то там из оксалоацетата синтезируется фосфоенолпируват. Далее из двух фосфоенолпируватов через серию реакций образуется одна молекула глюкозы.
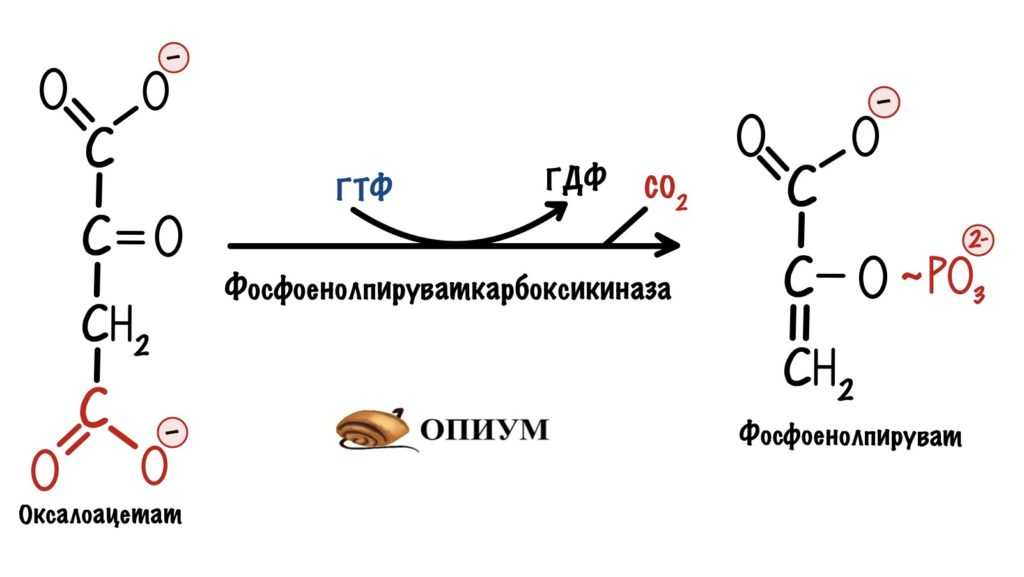 Синтез фосфоенолпирувата из оксалоацетата
Синтез фосфоенолпирувата из оксалоацетата
У фосфоенолпирувата есть и другой анаболический путь. Он может пойти по шагам глюконеогенеза, но остановиться на 3-фосфоглицерате. Из фосфоглицерата через серию реакций образуется серин. А из серина столько всего наворотить можно…
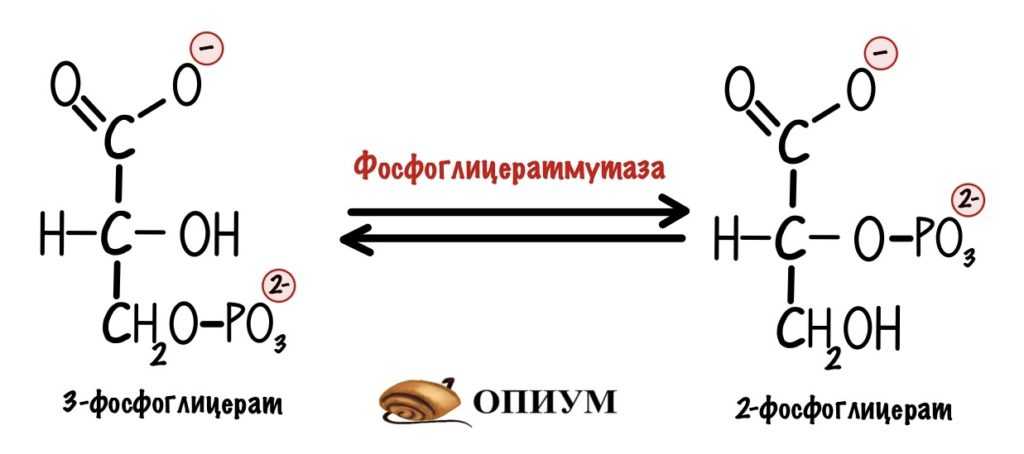
Образование 3-фосфоглицерата в глюконеогенезе
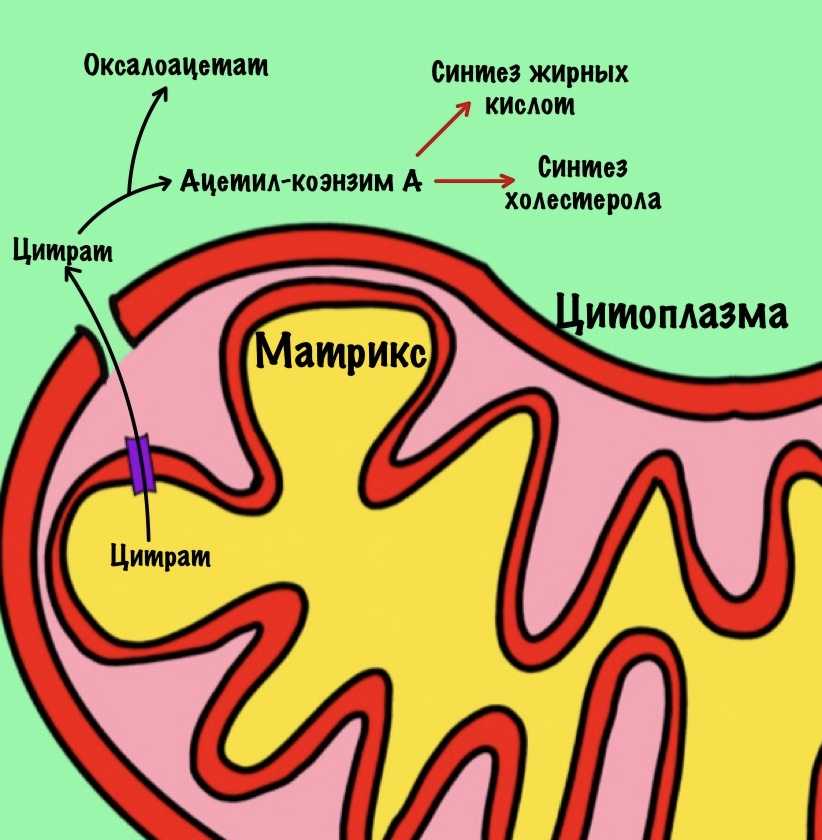
Цитрат. Для синтеза жиров и холестерола нужен ацетил-коэнзим А. Он находится в матриксе митохондрий, а ферменты биосинтеза в цитоплазме. Для ацетил-коэнзима А нет переносчиков во внутренней мембране митохондрий. А для цитрата есть, поэтому сначала ацетил-коэнзим А реагирует с оксалоацетатом. Далее цитрат переносится в цитоплазму, где из него образуются оксалоацетат и ацетил-коэнзим А. Ацетил-коэнзим А пойдет на синтез жирных кислот и холестерола. Из холестерола синтезируются стероидные гормоны.
Альфа-кетоглутарат. С помощью реакции трансаминирования из него образуется глутамат. Из глутамата можно получить гамма-аминомасляную кислоту (ГАМК), глутатион и глутамин. ГАМК — это нейромедиатор в ЦНС. Глутатион используется для реакций дезинтоксикации. А из глутамина синтезируются пурины.





Из глутамата синтезируются другие аминокислоты, например пролин и аргинин. Это опять-таки аминокислоты, использующиеся для синтеза белка.
Сукцинил-коэнзим А. Из сукцинил-коэнзима А синтезируются порфирины — входят в состав гема. Поэтому без него не было бы гемоглобина и миоглобина, а ещё цитохромов.
Вроде всё, но это в общих чертах. Пока что эти реакции вам не нужны, когда дойдёте до каждого вида обмена, то разберётесь с ними подробнее. Этот пунктик нужен был для того, чтобы показать глобальную роль ЦТК. Это не просто цикл для получения восстановленных коферментов НАДH и ФАДH2. Цитратный цикл — источник важнейших прекурсоров для многих видов обмена.
Регуляция общего пути катаболизма
Осталось поговорить про регуляцию работы пируватдегидрогеназного комплекса и ЦТК. Основная задача для регуляции — поддерживать энергетическое соотношение в клетке. Если в клетке достаточно АТФ, то общий путь катаболизма должен быть отключён. Если концентрация АТФ падает, а нарастает АДФ или АМФ, то нужно запускать общий путь катаболизма.
Основной механизм регуляции пируватдегидрогеназного комплекса и ЦТК — . Она достаточно быстро и чутко реагирует на изменения энергетического соотношения в клетке. Удивительно, что гормональной регуляции с помощью фосфорилирования/дефосфорилирования здесь нет.
Организм отдал регуляцию общего пути катаболизма на усмотрение клетки. Гликолиз и такого доверия не заслужили. С другой стороны, зачем регулировать каждый этап катаболизма гормонально? Достаточно установить контроль над специфическими путями катаболизма, но я отвлёкся.
Регуляция окислительного декарбоксилирования пирувата
Пируватдегидрогеназный комплекс состоит не только из трёх ферментов и пяти коферментов. С первым ферментом (пируватдегидрогеназой) связаны ещё две белковые субъединицы: киназа и фосфатаза. Киназа фосфорилирует пируватдегидрогеназу по -OH группе — переводит её в неактивную форму. Такое случается при избытке АТФ внутри митохондрии. Отдыхаем после тяжёлого дня, лёжа на диване. Фосфатаза же увеличивает свою работоспособность при снижении концентрации АТФ внутри клетки. Это приводит к дефосфорилированию пируватдегидрогеназы — активация фермента. Бежим за маршруткой — нужна энергия.
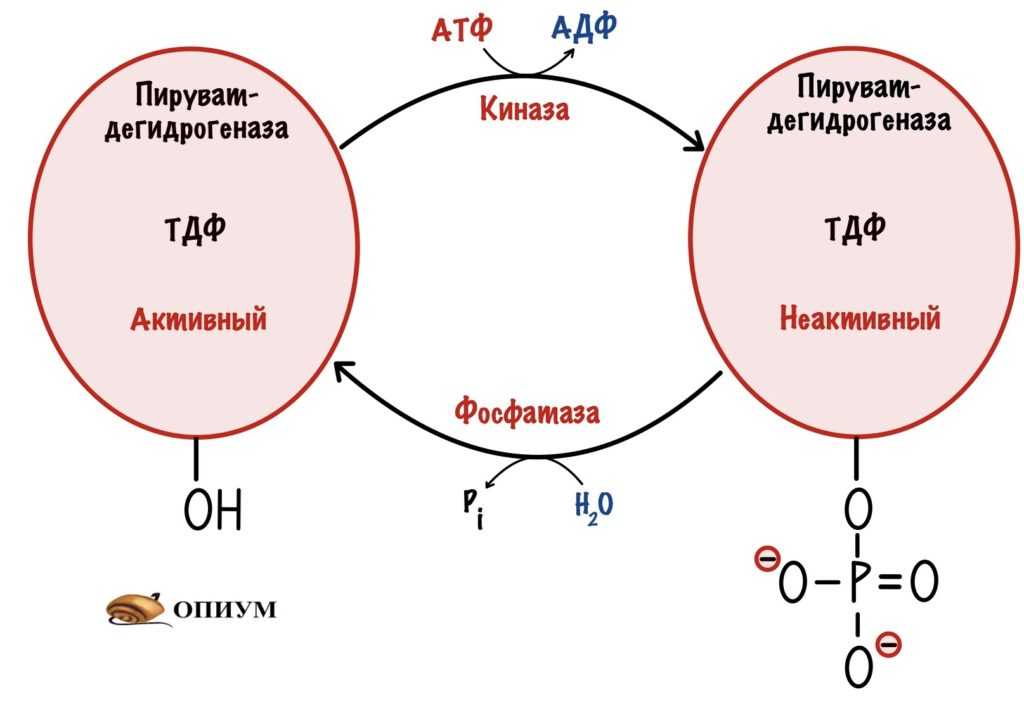 Киназа и фосфатаза пируватдегидрогеназного комплекса
Киназа и фосфатаза пируватдегидрогеназного комплекса
Это фофорилирование/дефосфорилирование, но не гормональное. Оно подкрепляет аллостерическую регуляцию. Киназа запускается при избытке АТФ, а фосфатаза при его дефиците. Но давайте перейдём к аллостерической регуляции.
Пируватдегидрогеназный комплекс аллостерически ингибируется: АТФ, НАДH, жирными кислотами, ацетил-коэнзимом А.
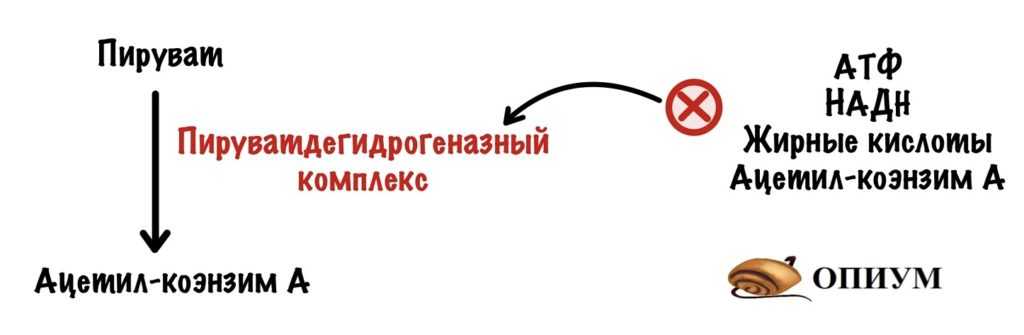 ПДК ингибируется: АТФ, НАДH, жирными кислотами и ацетил-коэнзимом А
ПДК ингибируется: АТФ, НАДH, жирными кислотами и ацетил-коэнзимом А
Все эти продукты накапливаются, когда клетке не нужна энергия. Понятно дело, что нужно остановить общий путь катаболизма, если в клетке куча АТФ и субстратов для её синтеза.
Пируватдегидрогеназный комплекс аллостерически активируется: АДФ, НАД+, коэнзимом А, кальцием.
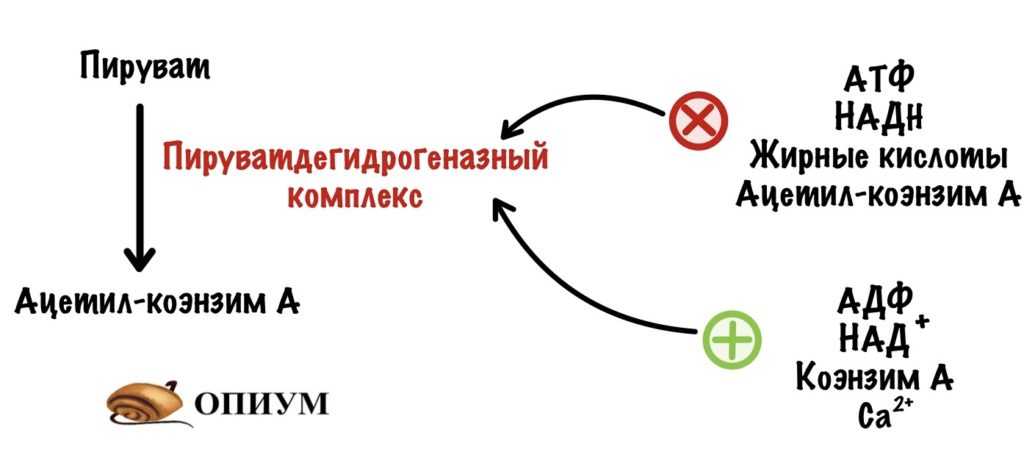 ПДК активируется: АДФ, НАД+, коэнзимом А и кальцием
ПДК активируется: АДФ, НАД+, коэнзимом А и кальцием
С первыми тремя всё ясно — они накапливаются, когда клетке нужна энергия, а её не хватает. Нужно ускорить работу пируватдегидрогеназного комплекса — образовать больше ацетил-коэнзима А. Чем больше ацетил-коэнзима А, тем больше энергии будет получено в ходе ЦТК. А что с кальцием? Кальций работает в мышцах и сердце. При сокращениях небольшая часть кальция попадает в митохондрию. Это стимулирует активность пируватдегидрогеназного комплекса и других ферментов ЦТК — об этом ниже.






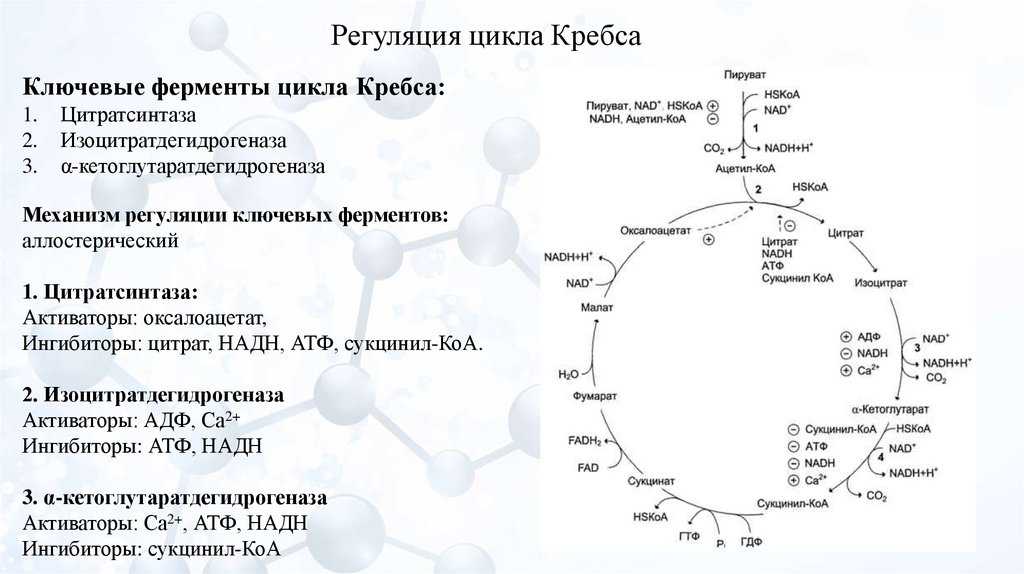
Регуляция цикла Кребса
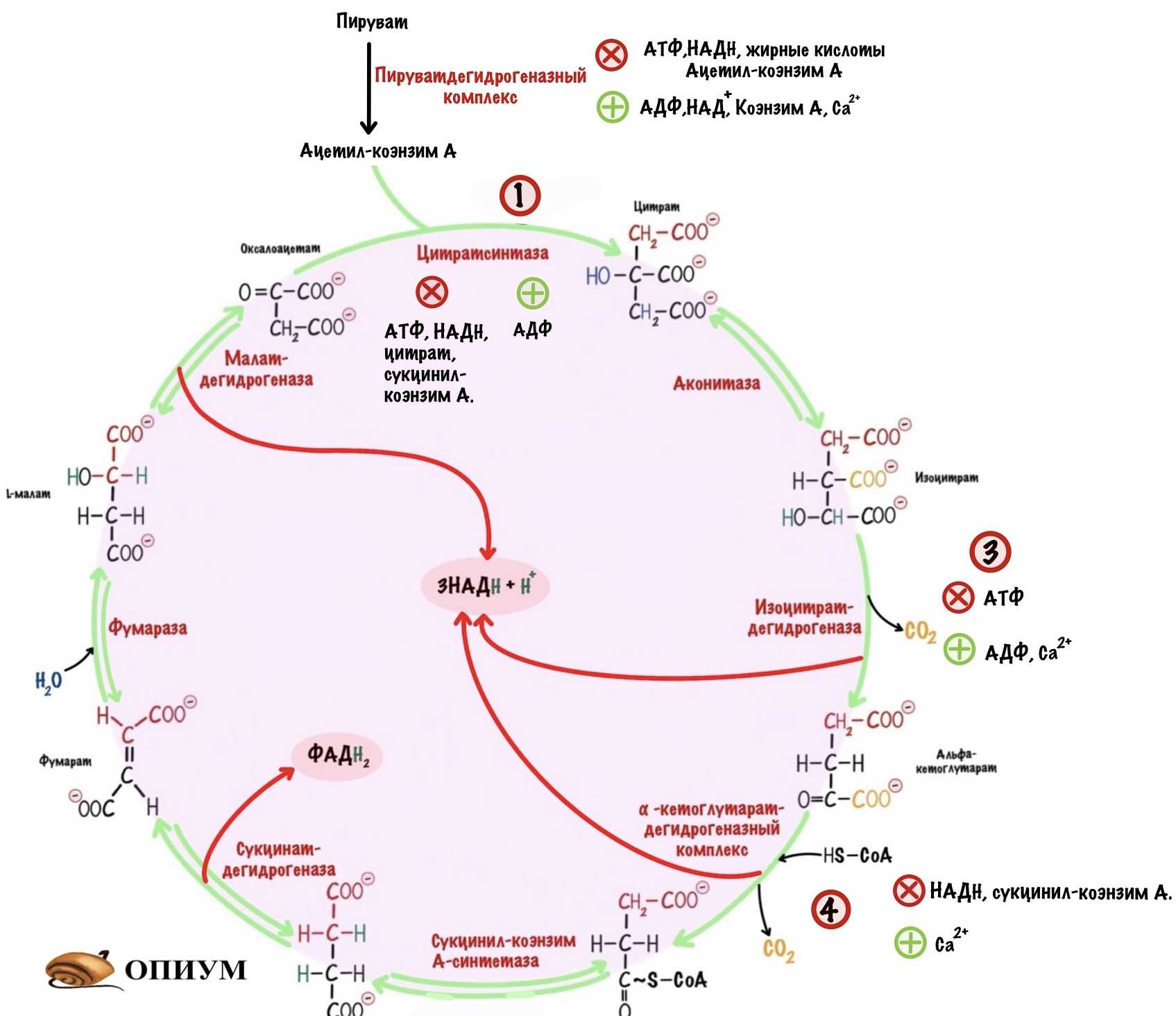
Идёт по трём необратимым реакциям: первой, третьей и четвёртой. Регулируются два фермента и один комплекс: цитратсинтаза, изоцитратдегидрогеназа и альфа-кетоглутаратдегидрогеназный комплекс.
Цитратсинтаза. Аллостерически ингибируется: АТФ, НАДH, цитратом и сукцинил-коэнзимом А. Активируется: АДФ.
Изоцитратдегидрогеназа. Ингибируется: АТФ. Активируется: АДФ и кальцием.
Альфа-кетоглутаратдегидрогеназный комплекс. Ингибируется: НАДH и сукцинил-коэнзим А. Активируется: кальцием.
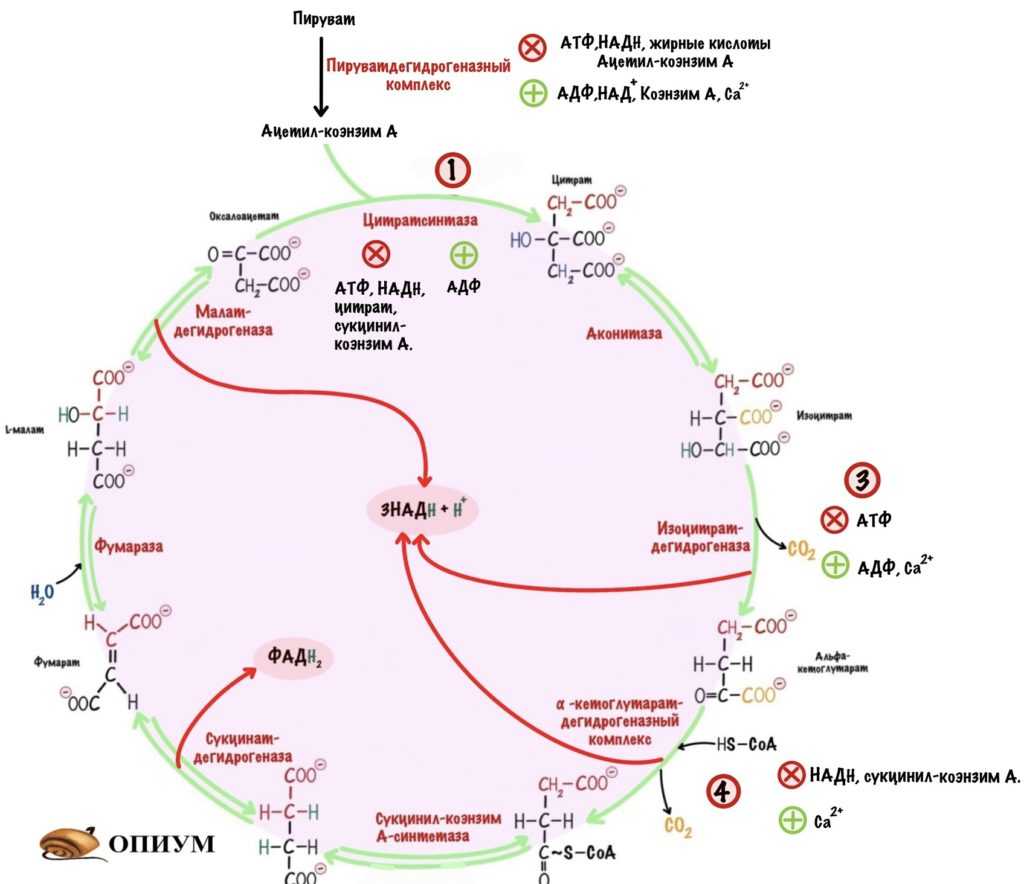 Регуляция ЦТК по первой, третьей и четвертой реакции.
Регуляция ЦТК по первой, третьей и четвертой реакции.
Видно, что наибольшее количество аллостерических регуляторов у цитратсинтазы и пируватдегидрогеназного комплекса. Это неудивительно, две эти реакции отвечают за старт цикла. Цитратсинтаза ингибируется не только продуктом своей реакции, но и продуктом четвертой реакции — сукцинил-коэнзимом А.