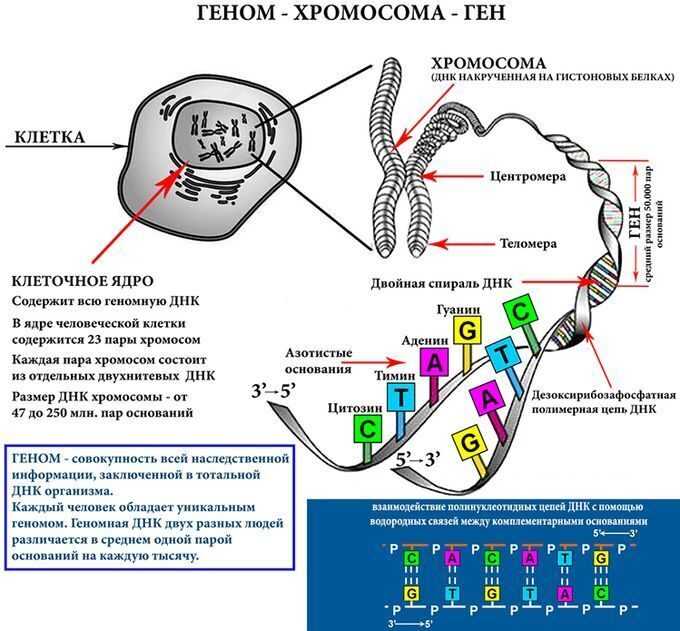
Строение ДНК
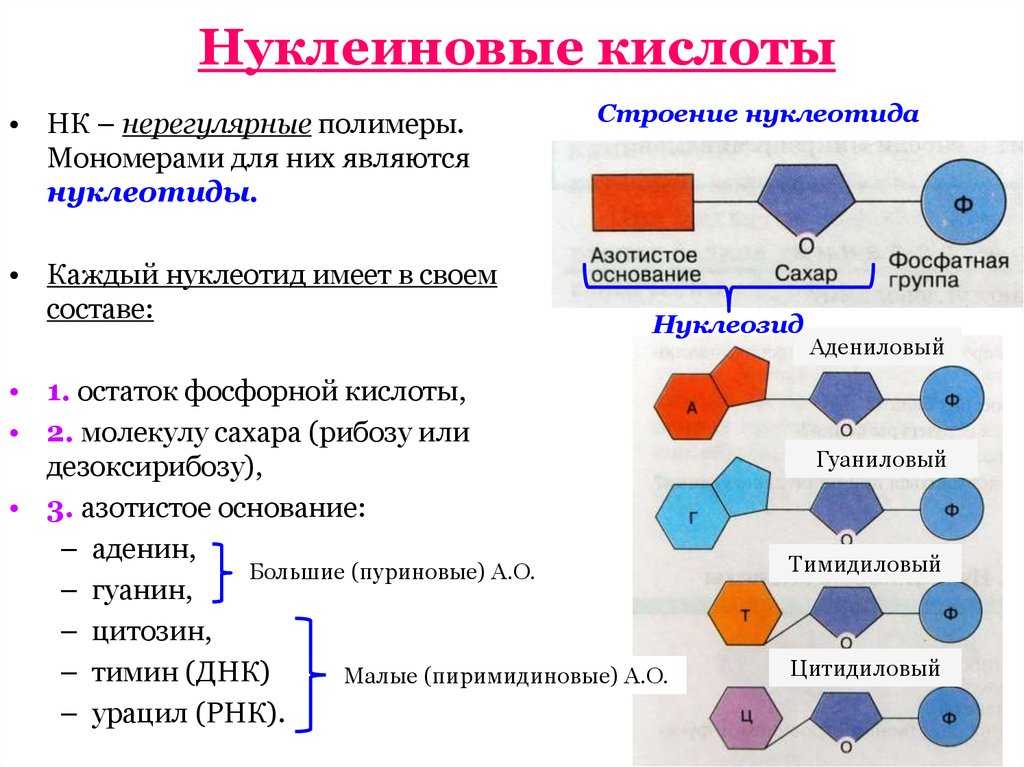
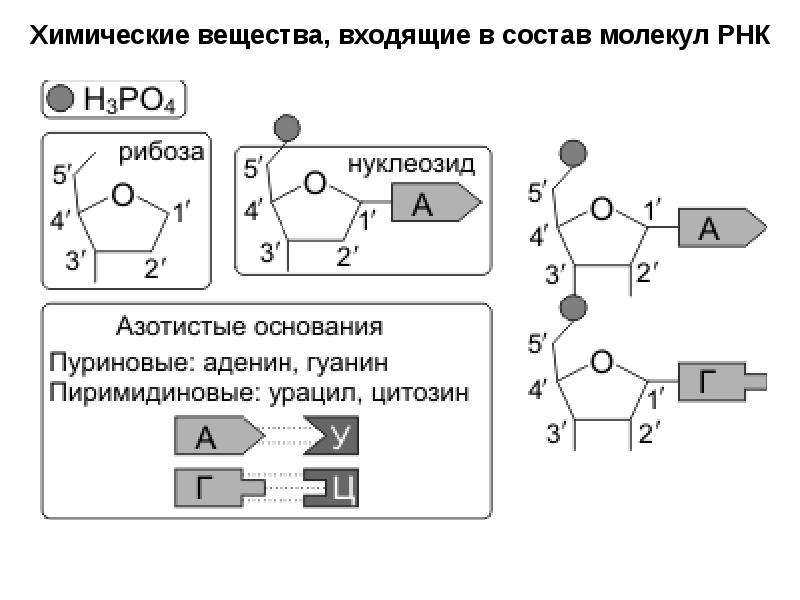
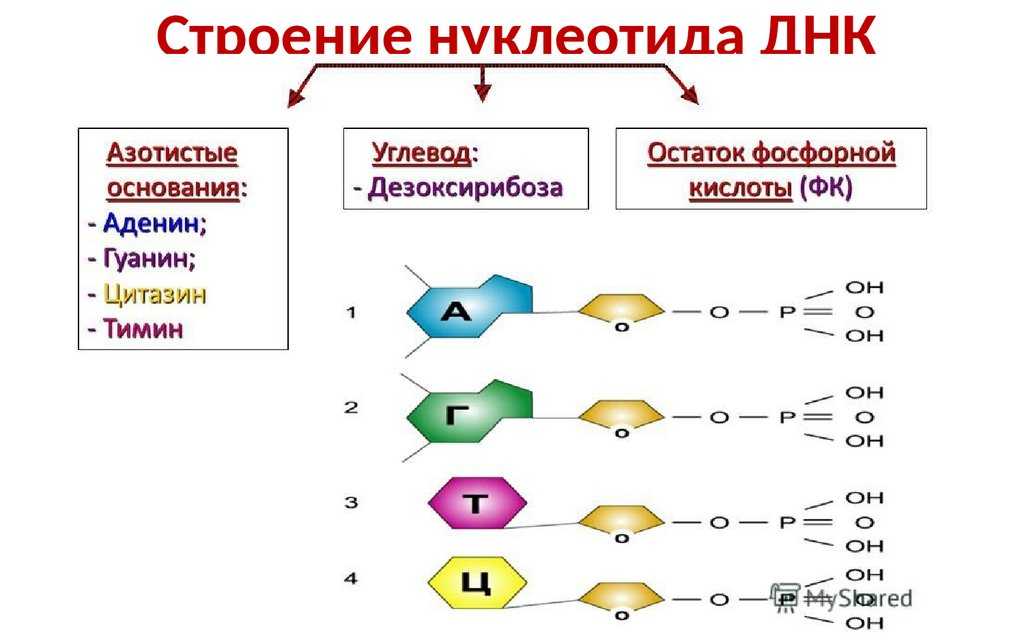
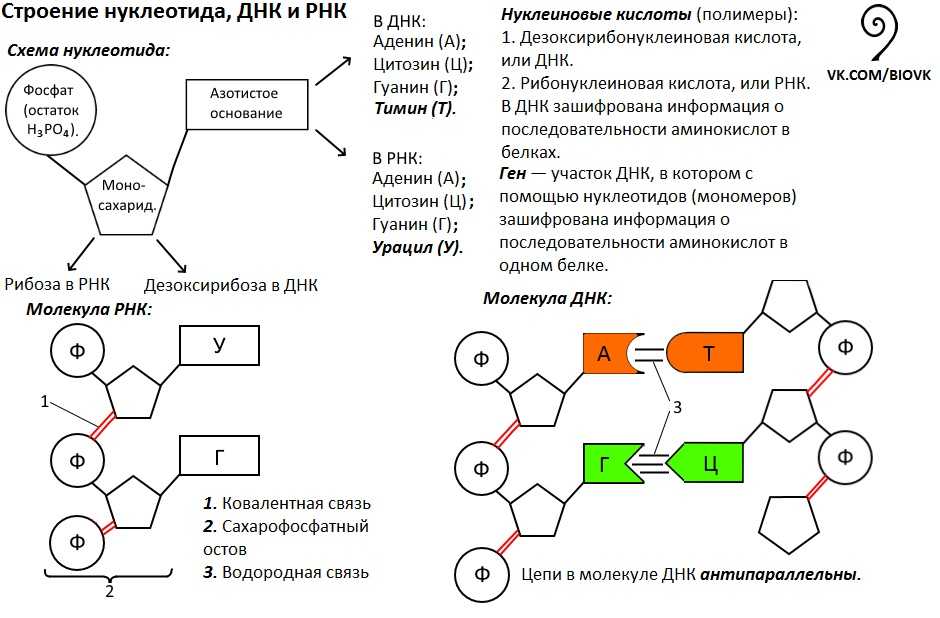
ДНК – это линейный органический полимер. Его мономерные звенья – нуклеотиды, которые, в свою очередь, состоят из:
- азотистого основания;
- пятиуглеродного сахара (пентозы);
-
фосфатной группы (рисунок 1).
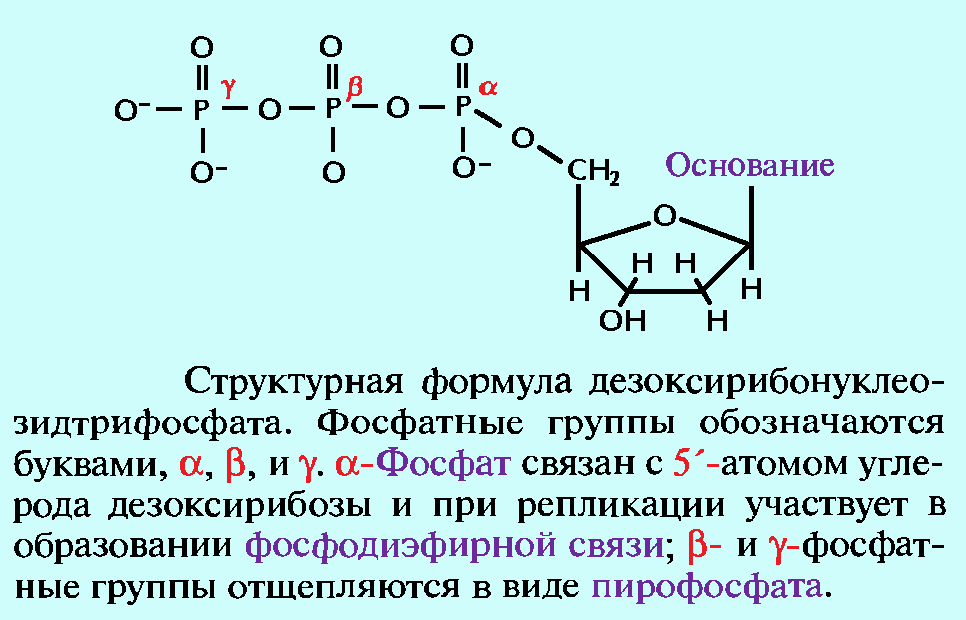
При этом, фосфатная группа присоединена к 5′-атому углерода моносахаридного остатка, а органическое основание — к 1′-атому.
Основания в ДНК бывают двух типов:
- Пуриновые: аденин ( А ) и гуанин (G);
-
Пиримидиновые: цитозин (С) и тимин (Т);(рисунок 2),
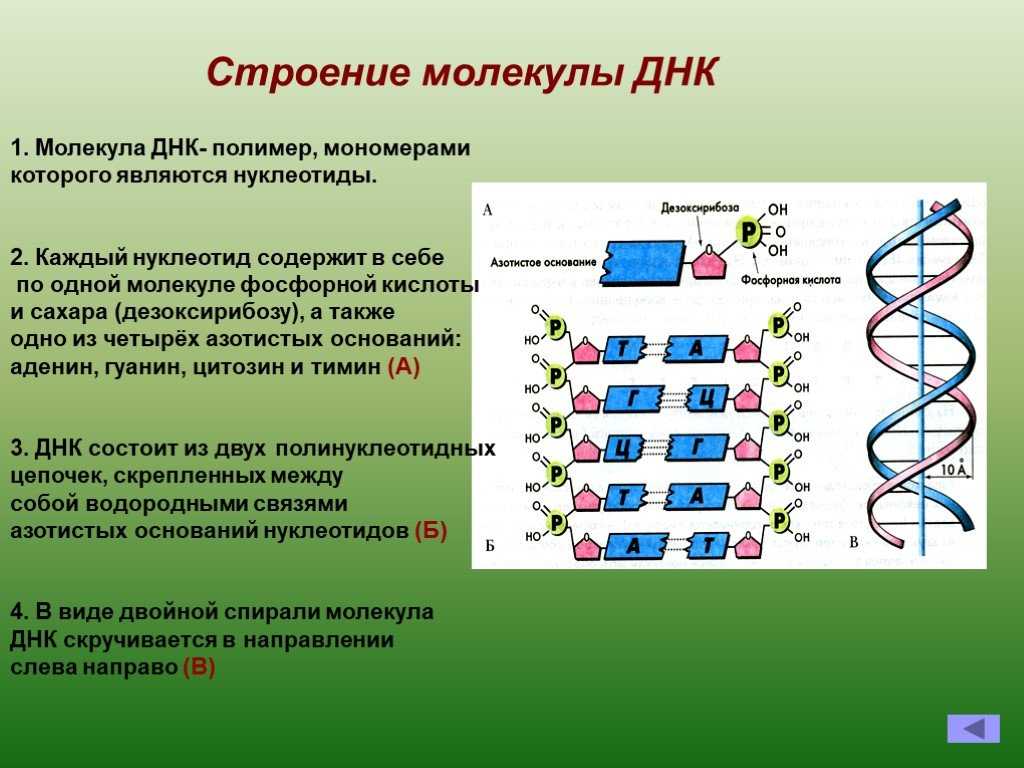
Строение нуклеотидов в молекуле ДНК
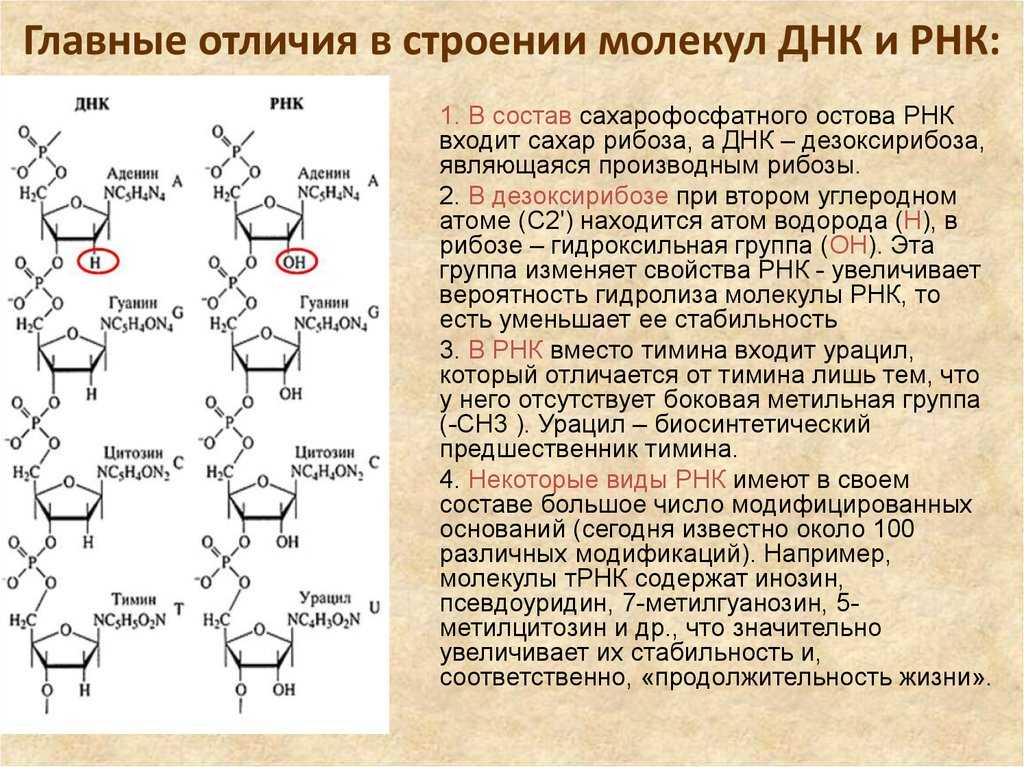
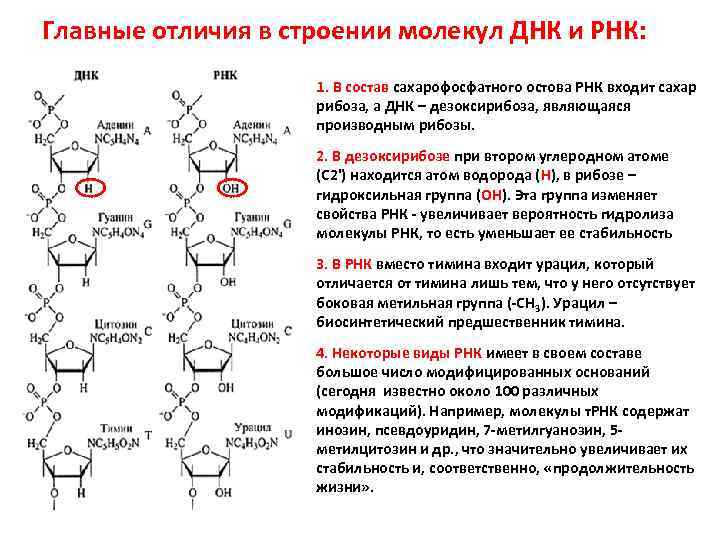
В ДНК моносахарид представлен 2′-дезоксирибозой, содержащей только 1 гидроксильную группу (ОН), а в РНК — рибозой, имеющей 2 гидроксильные группы (OH).
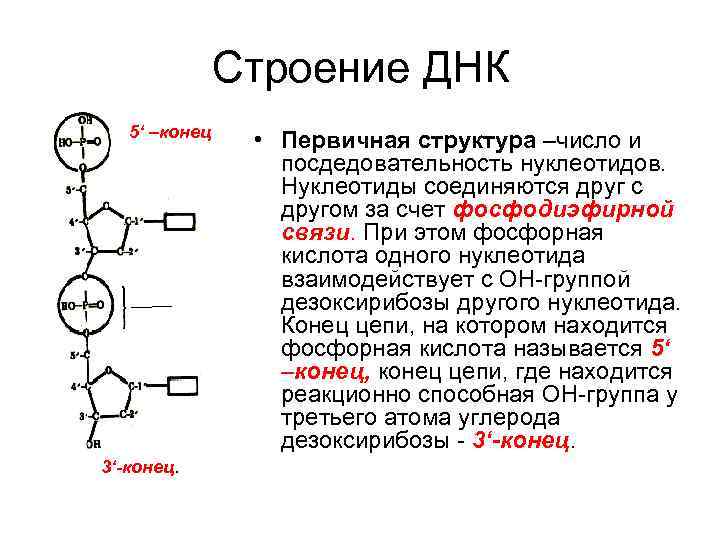
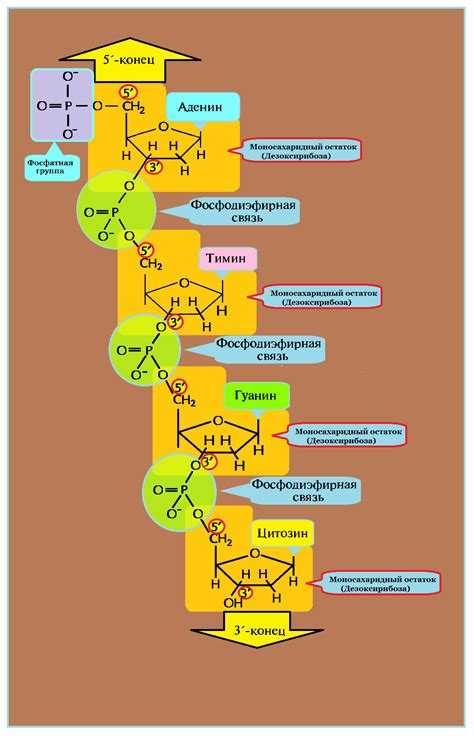
Нуклеотиды соединены друг с другом фосфодиэфирными связями, при этом фосфатная группа 5′-углеродного атома одного нуклеотида связана с З’-ОН-группой дезоксирибозы соседнего нуклеотида (рисунок 1). На одном конце полинуклеотидной цепи находится З’-ОН-группа (З’-конец), а на другом — 5′-фосфатная группа (5′-конец).
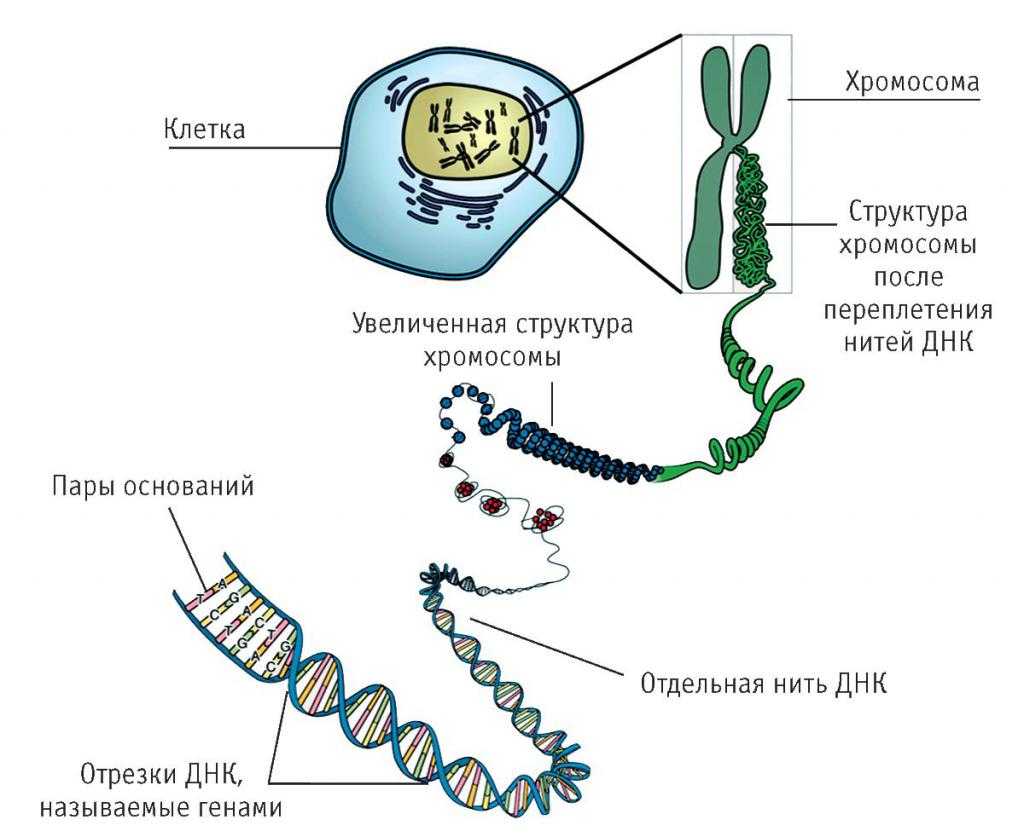
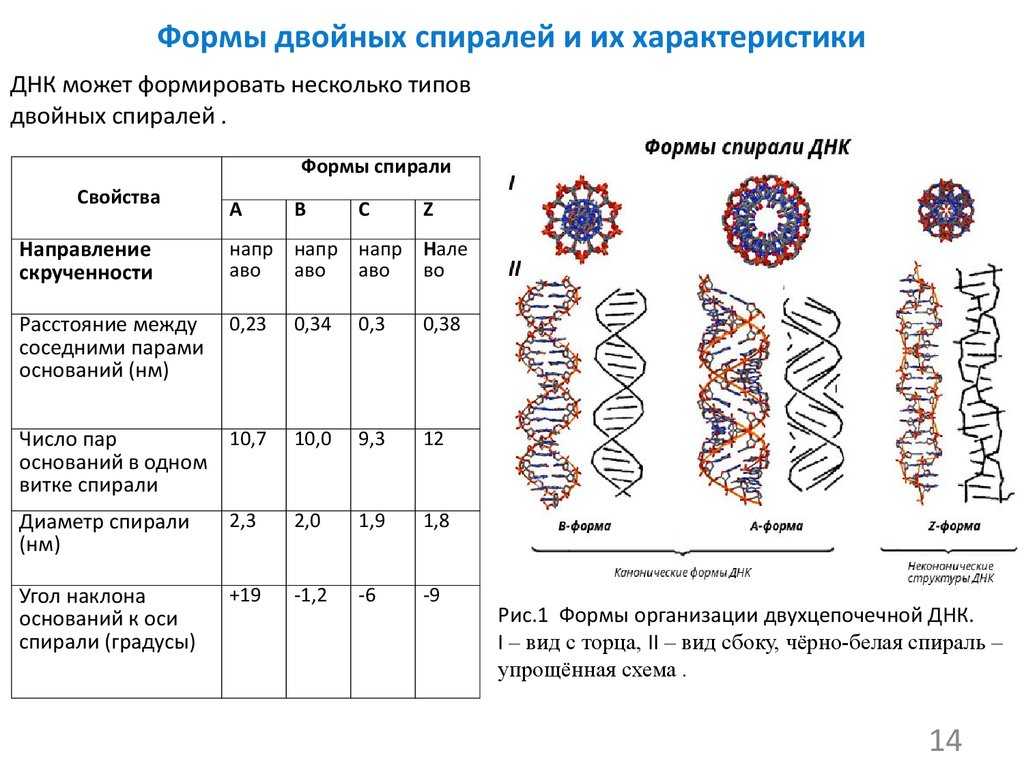
Уровни структуры ДНК
Принято выделять 3 уровня структуры ДНК:
- первичную;
- вторичную;
- третичную.
Первичная структура ДНК – это последовательность расположения нуклеотидов в полинуклеотидной цепи ДНК.
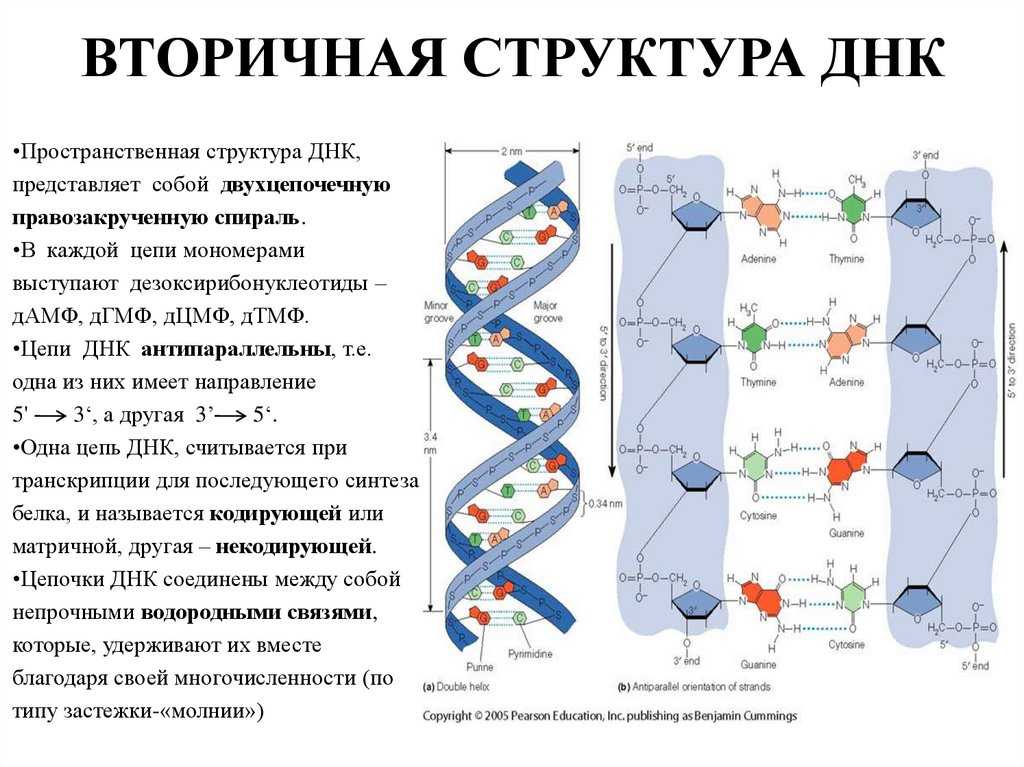
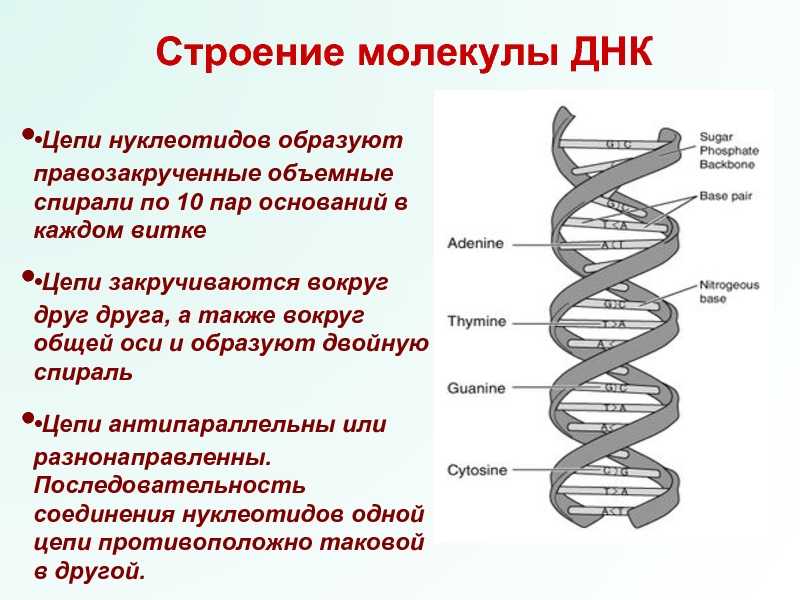
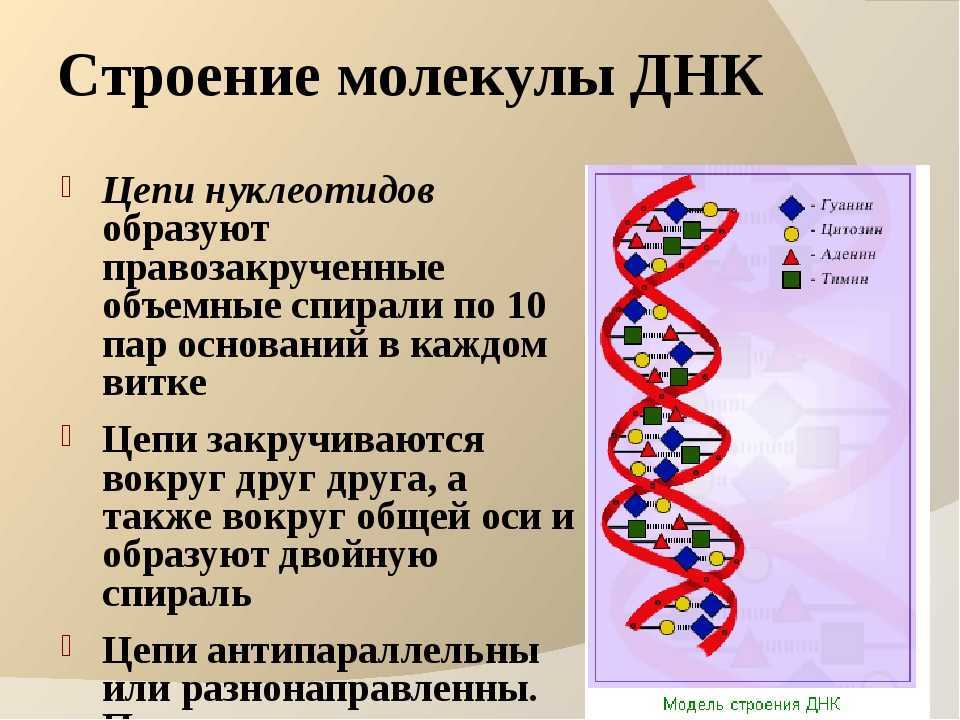
Вторичная структура ДНК стабилизируется водородными связями между комплементарными парами оснований и представляет собой двойную спираль из двух антипараллелных цепочек, закрученных вправо вокруг одной оси.
Общий виток спирали- 3,4нм, расстояние между цепочками 2нм.
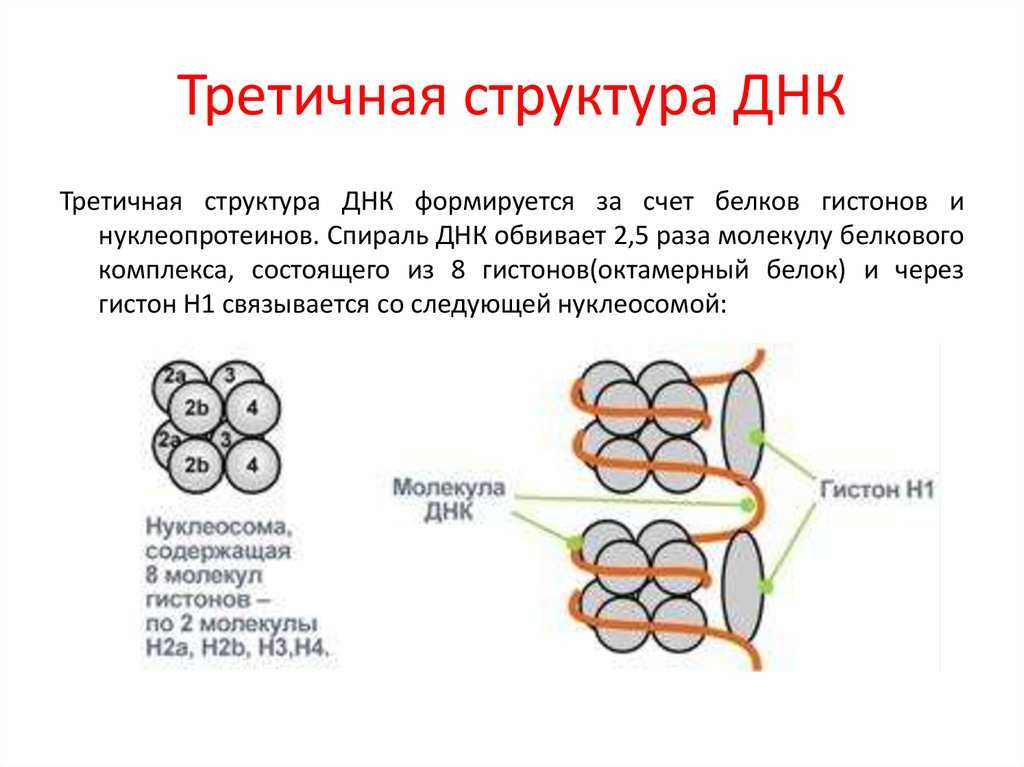
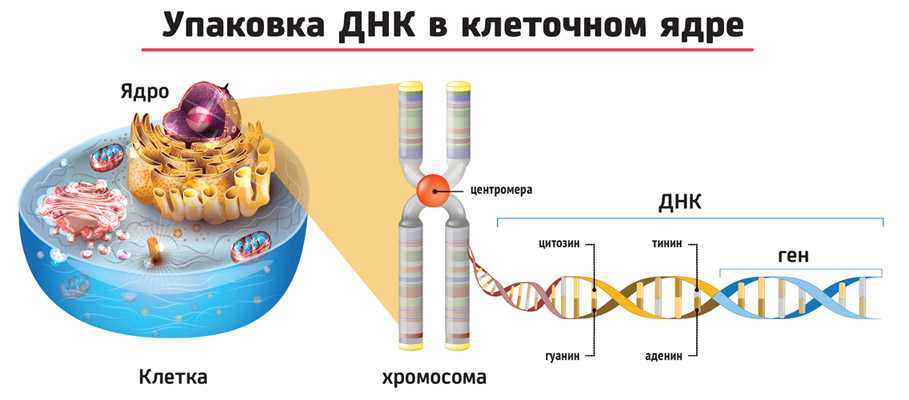
Третичная структура ДНК – суперсперализация ДНК. Двойная спираль ДНК на некоторых участках может подвергаться дальнейшей спирализации с образованием суперспирали или открытой кольцевой формы, что часто вызвано ковалентным соединением их открытых концов. Суперспиральная структура ДНК обеспечивает экономную упаковку очень длинной молекулы ДНК в хромосоме. Так, в вытянутой форме длина молекулы ДНК составляет 8 см, а в форме суперспирали укладывается в 5 нм.
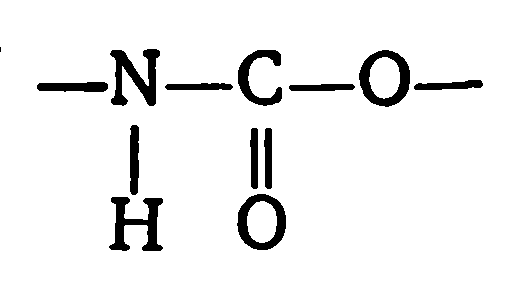


Правило Чаргаффа
Правило Э. Чаргаффа – это закономерность количественного содержания азотистых оснований в молекуле ДНК:
- У ДНК молярные доли пуриновых и пиримидиновых оснований равны: А+ G = C + Т или (А + G)/(C + Т)=1.
- В ДНК количество оснований с аминогруппами (А +C) равно количеству оснований с кетогруппами (G + Т): А +C= G + Т или (А +C)/(G + Т)= 1
- Правило эквивалентности, то есть : А=Т, Г=Ц; А/Т = 1; Г/Ц=1.
- Нуклеотидный состав ДНК у организмов различных групп специфичен и характеризуется коэффициентом специфичности: (Г+Ц)/(А+Т). У высших растений и животных коэффициент специфичности меньше 1, и колеблется незначительно: от 0,54 до 0,98, у микроорганизмов он больше 1.
Модель ДНК Уотсона-Крика
Б 1953 г. Джеймс Уотсон и Фрэнсис Крик, основываясь на данных рентгеноструктурного анализа кристаллов ДНК, пришли к выводу, что нативная ДНК состоит из двух полимерных цепей, образующих двойную спираль (рисунок 3).
Навитые одна на другую полинуклеотидные цепи удерживаются вместе водородными связями, образующимися между комплементарными основаниями противоположных цепей (рисунок 3). При этом аденин образует пару только с тимином, а гуанин — с цитозином. Пара оснований А—Т стабилизируется двумя водородными связями, а пара G—С — тремя.
Длина двухцепочечной ДНК обычно измеряется числом пар комплементарных нуклеотидов (п.н.). Для молекул ДНК, состоящих из тысяч или миллионов пар нуклеотидов, приняты единицы т.п.н. и м.п.н. соответственно. Например, ДНК хромосомы 1 человека представляет собой одну двойную спираль длиной 263 м.п.н.
Сахарофосфатный остов молекулы, который состоит из фосфатных групп и дезоксирибозных остатков, соединенных 5’—З’-фосфодиэфирными связями, образует «боковины винтовой лестницы», а пары оснований А—Т и G—С — ее ступеньки (рисунок 3).
Рисунок 3: Модель ДНК Уотсона-Крика
Цепи молекулы ДНК антипараллельны: одна из них имеет направление 3’→5′, другая 5’→3′. В соответствии с принципом комплементарности, если в одной из цепей имеется нуклеотидная последовательность 5-TAGGCAT-3′, то в комплементарной цепи в этом месте должна находиться последовательность 3′-ATCCGTA-5′. В этом случае двухцепочечная форма будет выглядеть следующим образом:


- 5′-TAGGCAT-3′
- 3-ATCCGTA-5′.
В такой записи 5′-конец верхней цепи всегда располагают слева, а 3′-конец — справа.
Носитель генетической информации должен удовлетворять двум основным требованиям: воспроизводиться (реплицироваться) с высокой точностью и детерминировать (кодировать) синтез белковых молекул.
Модель ДНК Уотсона—Крика полностью отвечает этим требованиям, так как:
- согласно принципу комплементарности каждая цепь ДНК может служить матрицей для образования новой комплементарной цепи. Следовательно, после одного раунда репликации образуются две дочерние молекулы, каждая из которых имеет такую же нуклеотидную последовательность, как исходная молекула ДНК.
- нуклеотидная последовательность структурного гена однозначно задает аминокислотную последовательность кодируемого ею белка.