
Потенциал воображения
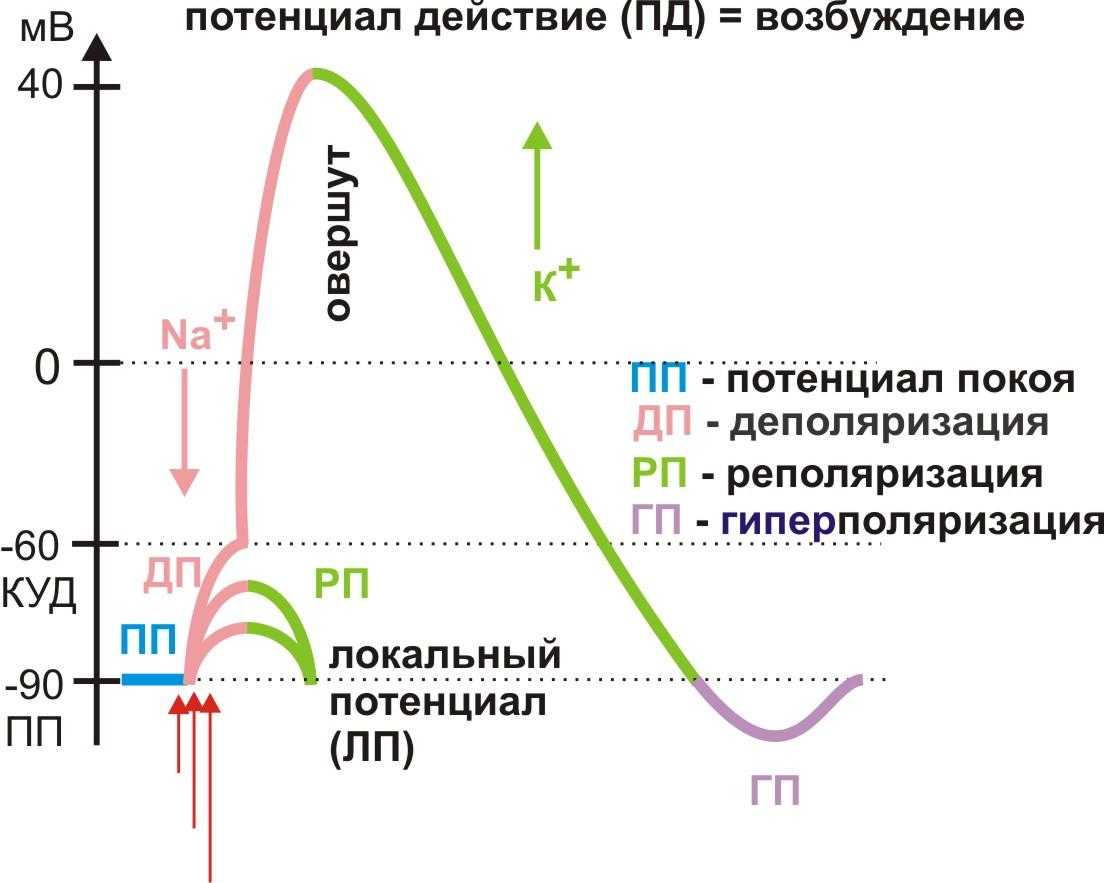
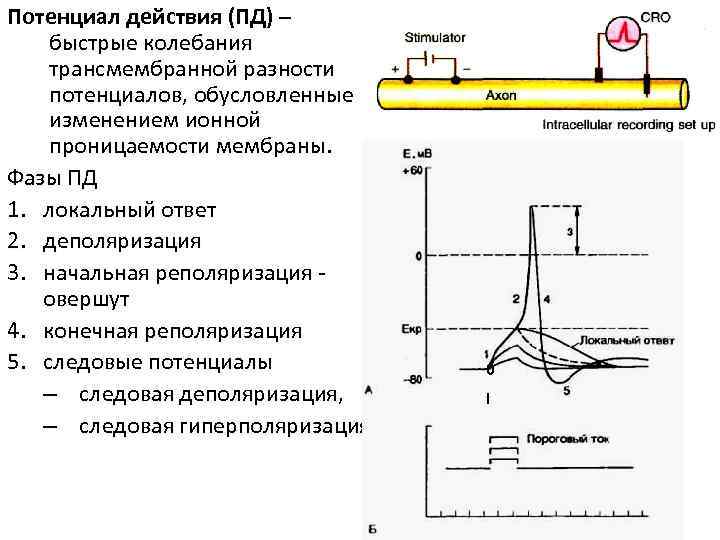
Какие же условия нужны для того, чтобы появился потенциал действия? Прежде всего, должно «прилететь» раздражение либо извне, либо благодаря блуждающим по мембране локальным потенциалам, которые, объединившись на аксональном холмике (его можно даже назвать «родильным домом»), переходят в реальный потенциал действия. С этого момента он становится нервным импульсом и передаётся по аксону, вызывая в синаптическом окончании выброс нейромедиаторов, которые устремляются к мембране другой клетки через синаптическую щель (узнаёте картину, описанную в начале статьи?)
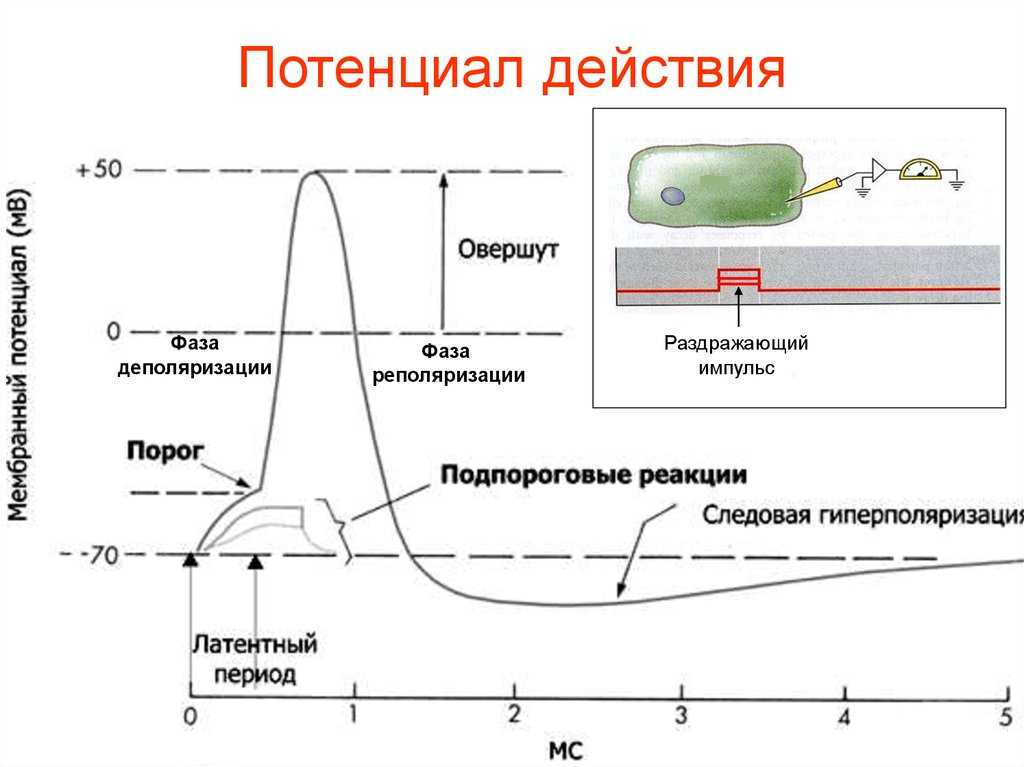
Кроме того, то самое раздражение должно быть достаточным по силе, чтобы достичь порога возбуждения, достаточным по времени действия и скорости нарастания. Имеет значение даже его направление (если оно электрическое, то в зависимости от полюса направление будет либо в сторону деполяризации, либо в сторону гиперполяризации).
Так, стоп. А почему нас это волнует? Поясним: чем мембрана более деполяризована, тем клетка более активна и готова на ответ в виде потенциала действия. Например, если её начальный заряд не -70 мВ, а -60. Именно поэтому любое малейшее движение (зачёркнуто) раздражение, даже подпороговой силы, будет способно клетку возбудить и заставить её «вздрогнуть в конвульсии».
Когда же она находится в состоянии гиперполяризации, ей глубоко «всё равно», какими пороговыми раздражителями на неё воздействуют. Например тогда, когда мембранный потенциал составляет -100мВ, и до порога нужно преодолеть целых 45мВ. Она «не в духе», и только сила гораздо выше обычного для неё порога сможет «достать» её из этого состояния и помочь ответить потенциалом действия. Но уж как только она достигает этого порогового значения, то отвечает со всей той силой и «пылкостью», на которую способна (согласно закону «всё или ничего»). А измеряется уровень «пылкости» количеством имеющихся в мембране потенциал-зависимых Na-каналов, которые могут открыться одновременно.

Движение ионов через каналы в процессе распространения потенциала действия
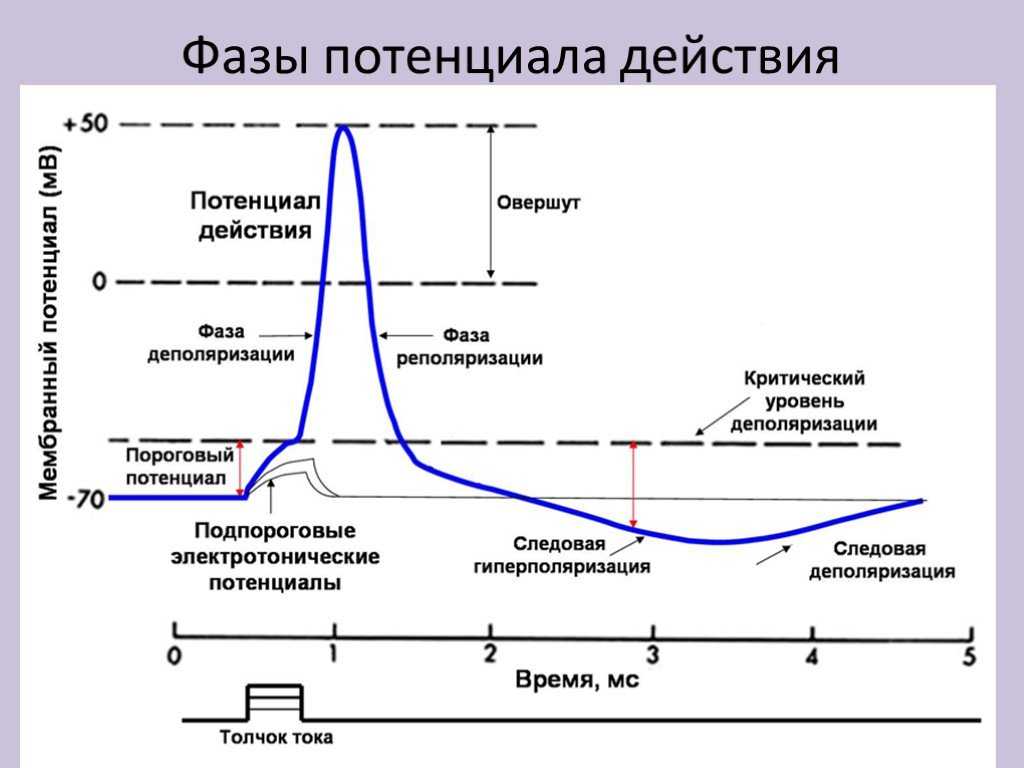
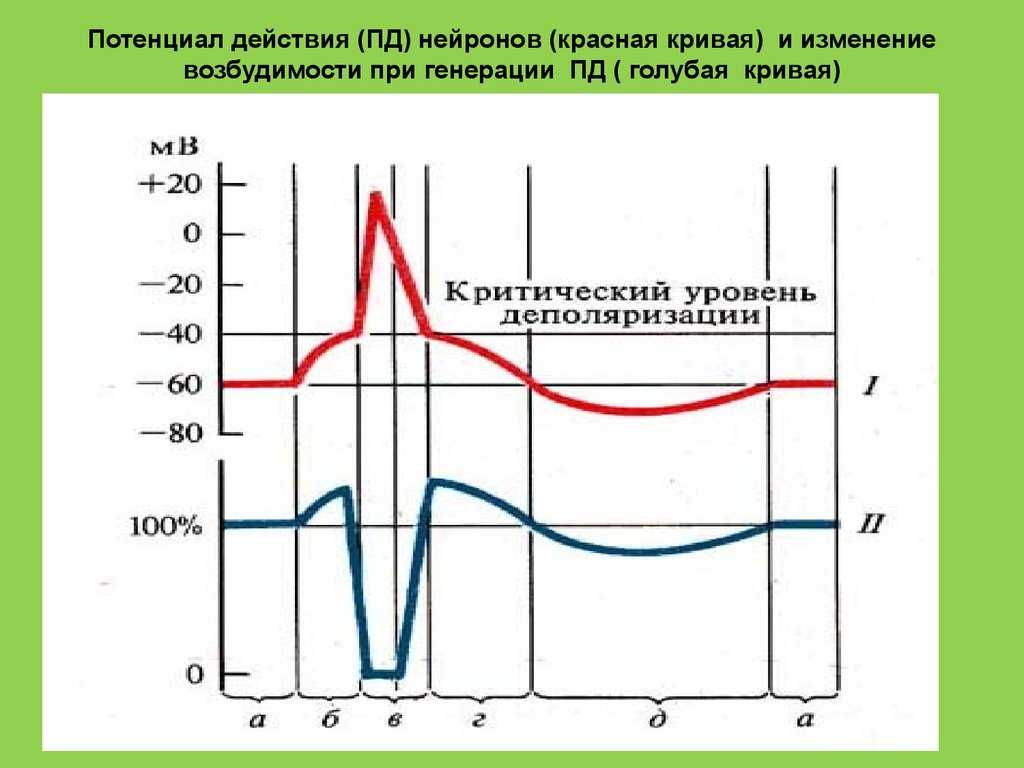
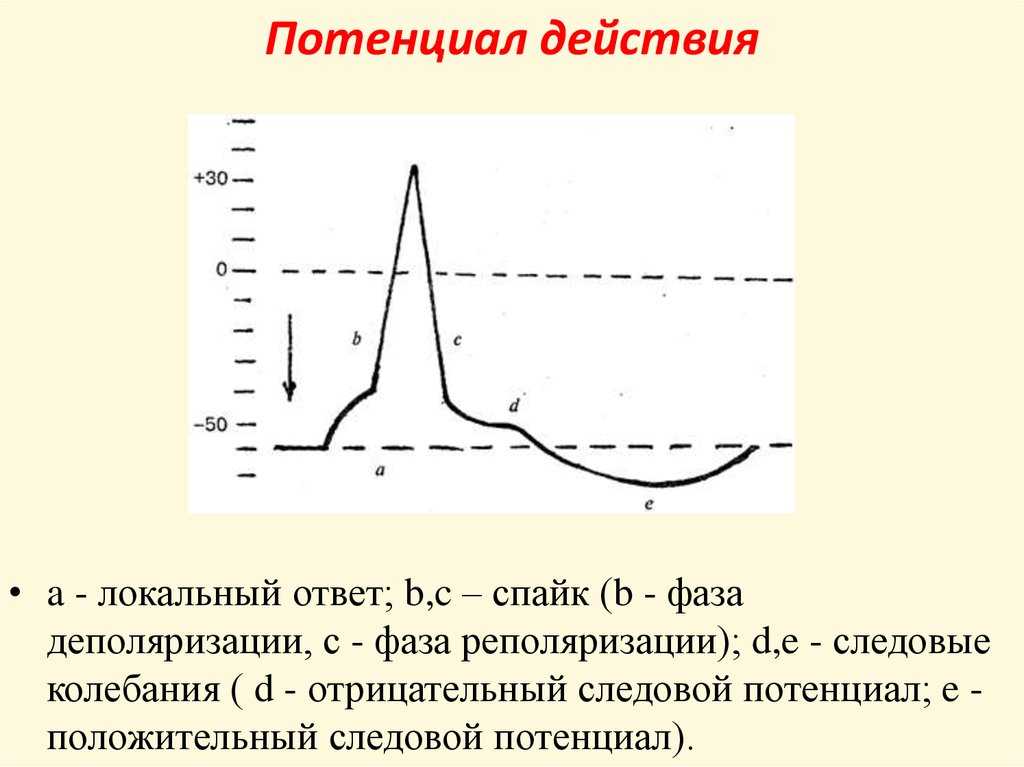
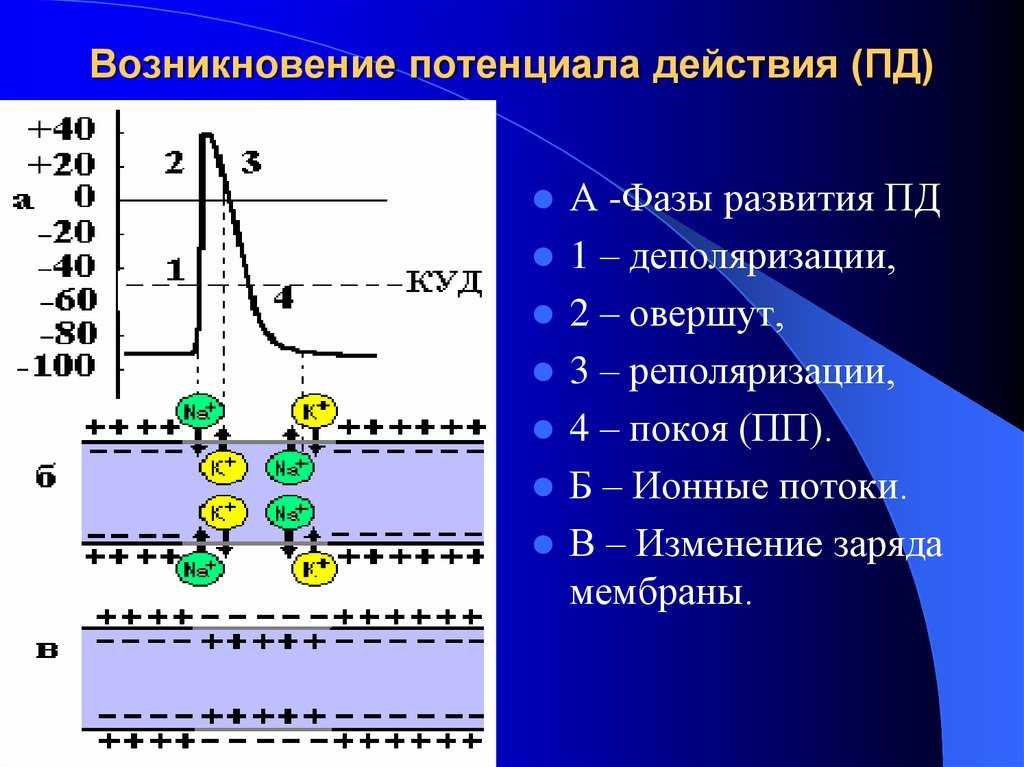
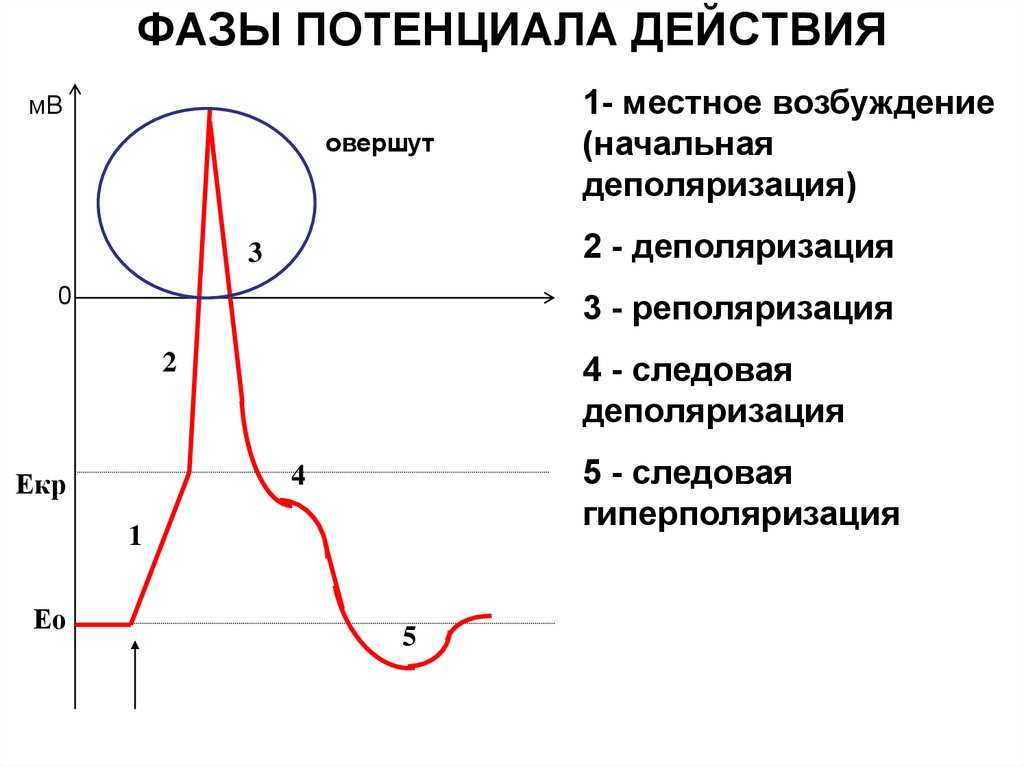
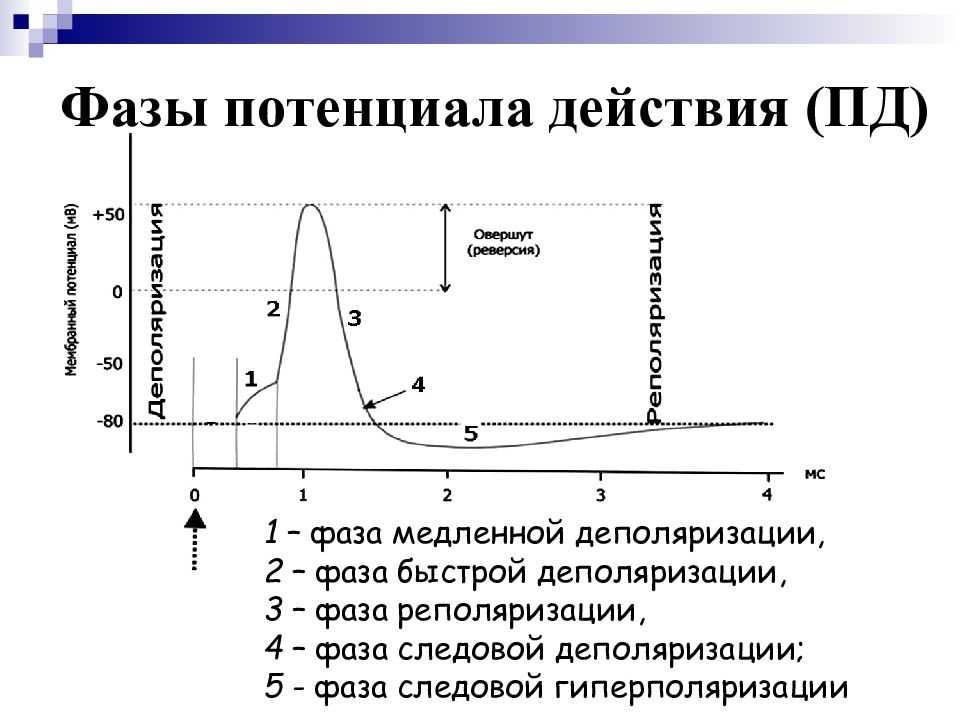
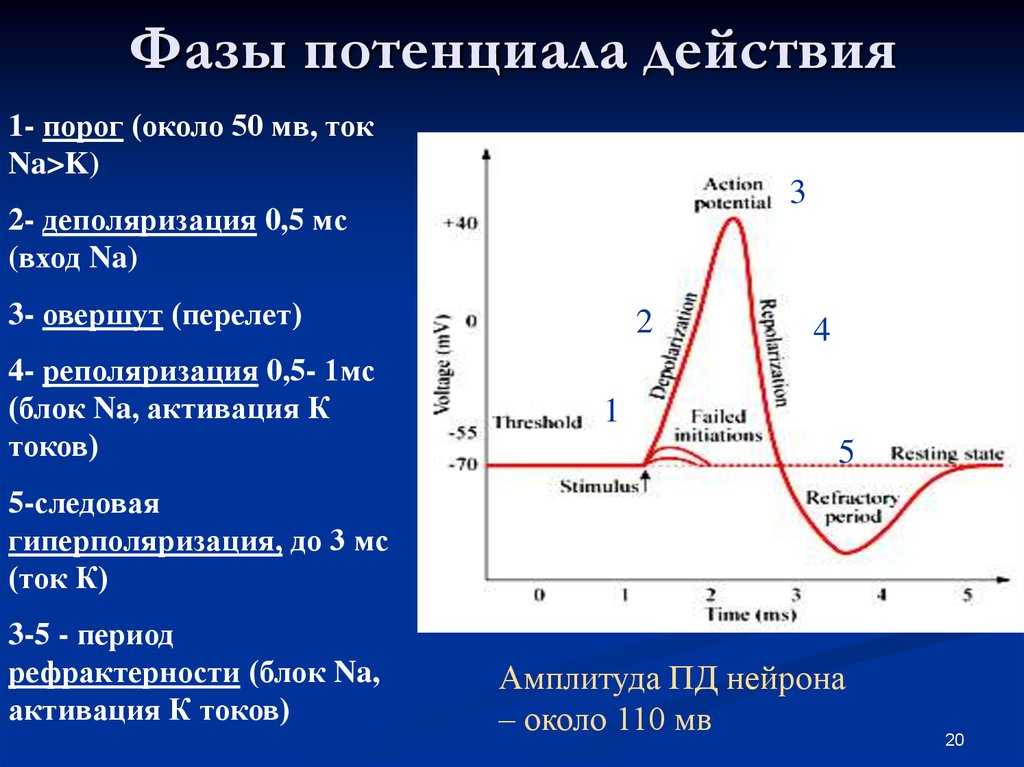
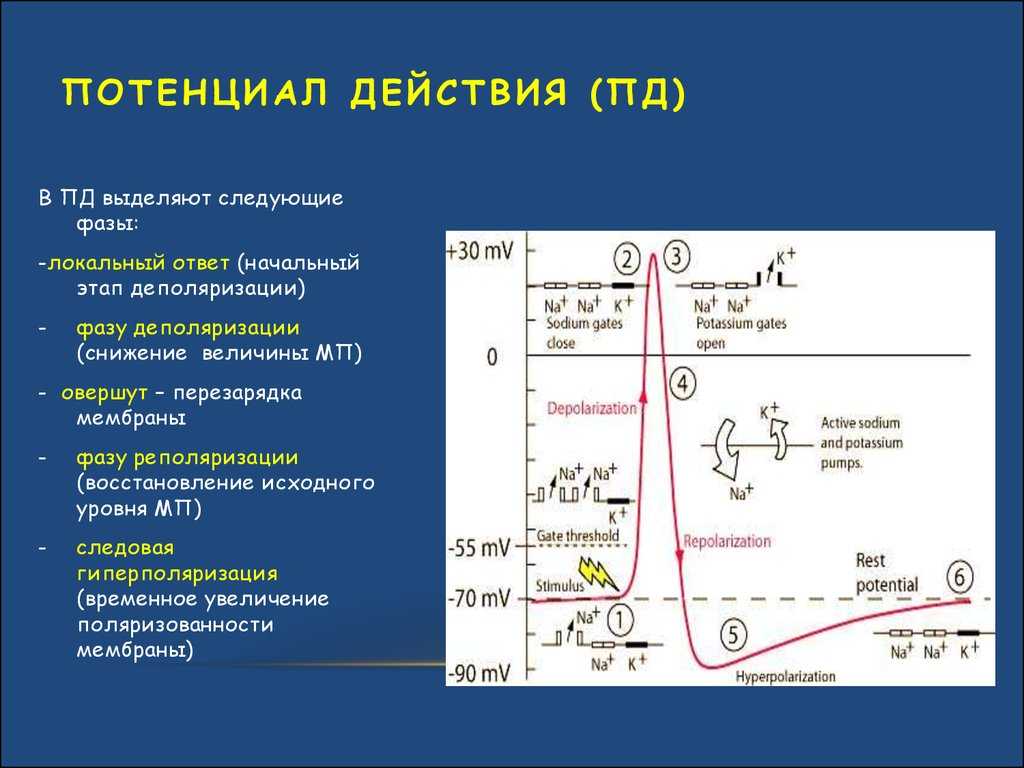
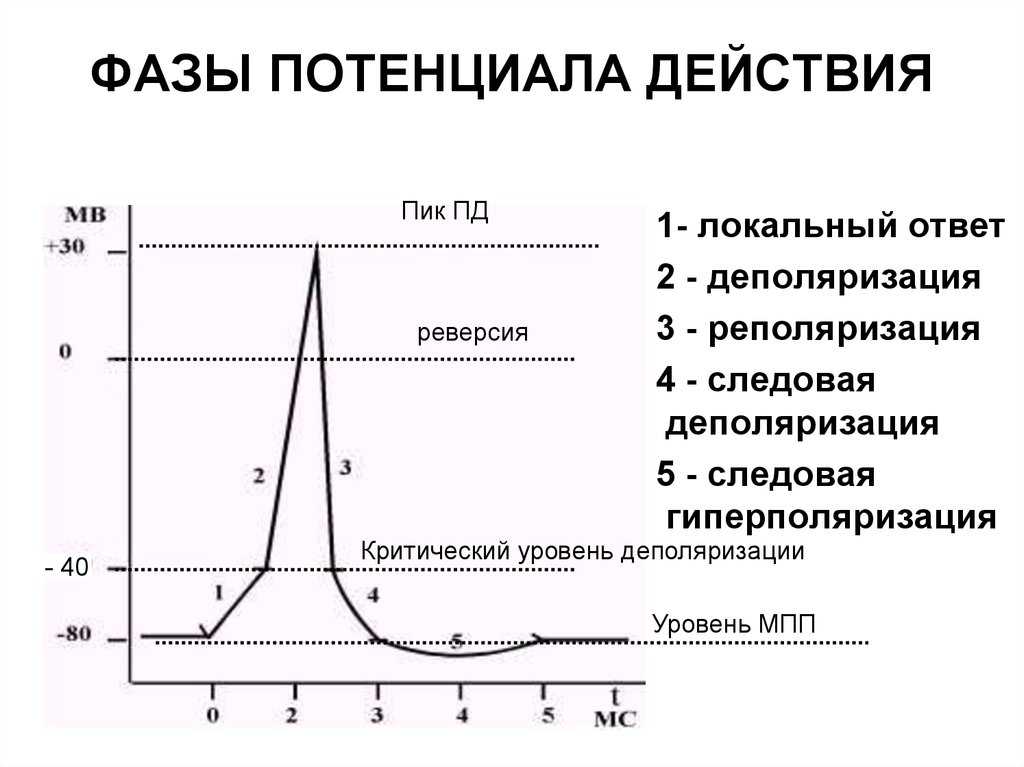
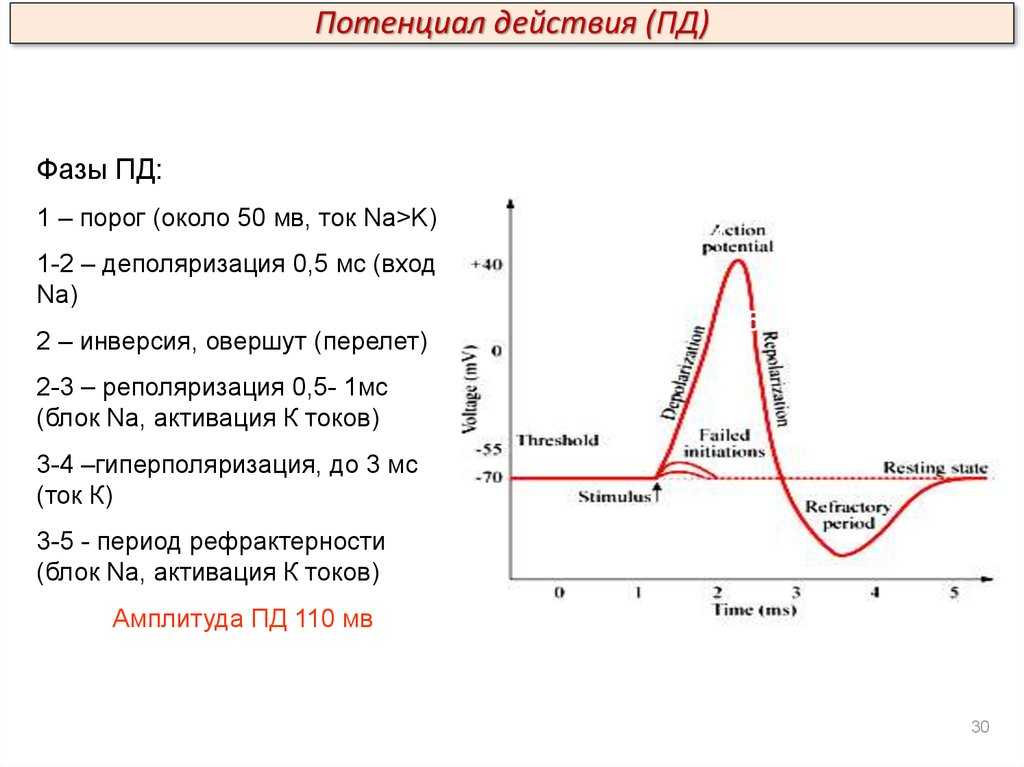
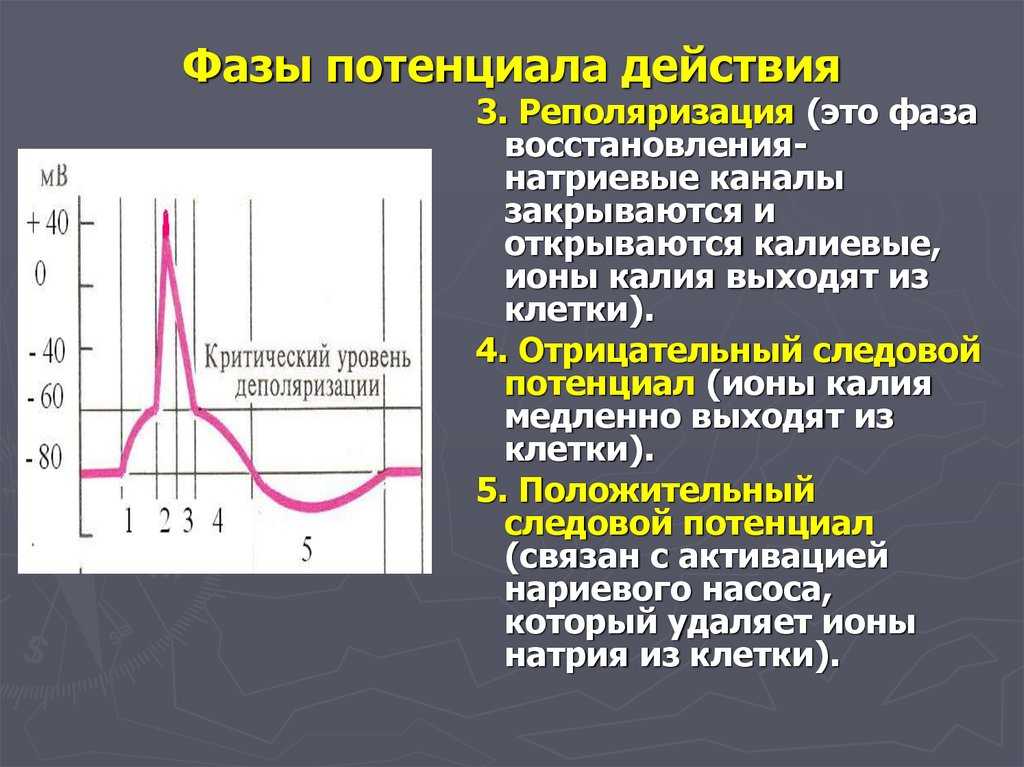
Но и это ещё не всё. Оказывается, на разных стадиях развернувшегося потенциала действия клетка тоже поддаётся (или не поддаётся) влиянию. Если попытаться её тронуть на этапе быстрой деполяризации и сразу после пика, то ничего не произойдёт, потому что в первом случае все возможные Na-каналы уже будут открыты, а во втором – инактивированы (закрыты на обе «щеколды»). Этот период называется периодом абсолютной рефрактерности. Но если ещё немного подождать и попытаться пораздражать мембрану за овершутом, то мы увидим ответ, хотя он будет достаточно слабым – период относительной рефрактерности. Это потому, что некоторые натриевые каналы уже «оправятся» и наполовину откроются, готовясь к новой серии возбуждения. Сразу за ним идёт короткий период повышенной возбудимости, так как клетка ещё не «остыла» и готова откликнуться потенциалом на любой слабый сигнал.
Если же мы потерпим ещё немного, избежав период гиперполяризации, и дадим раздражение, то увидим, что мембрана снова гипервозбудима, потому что попадём в так называемый следовый потенциал.
Такие вот клеточные «американские горки».
Ближе к «телу»
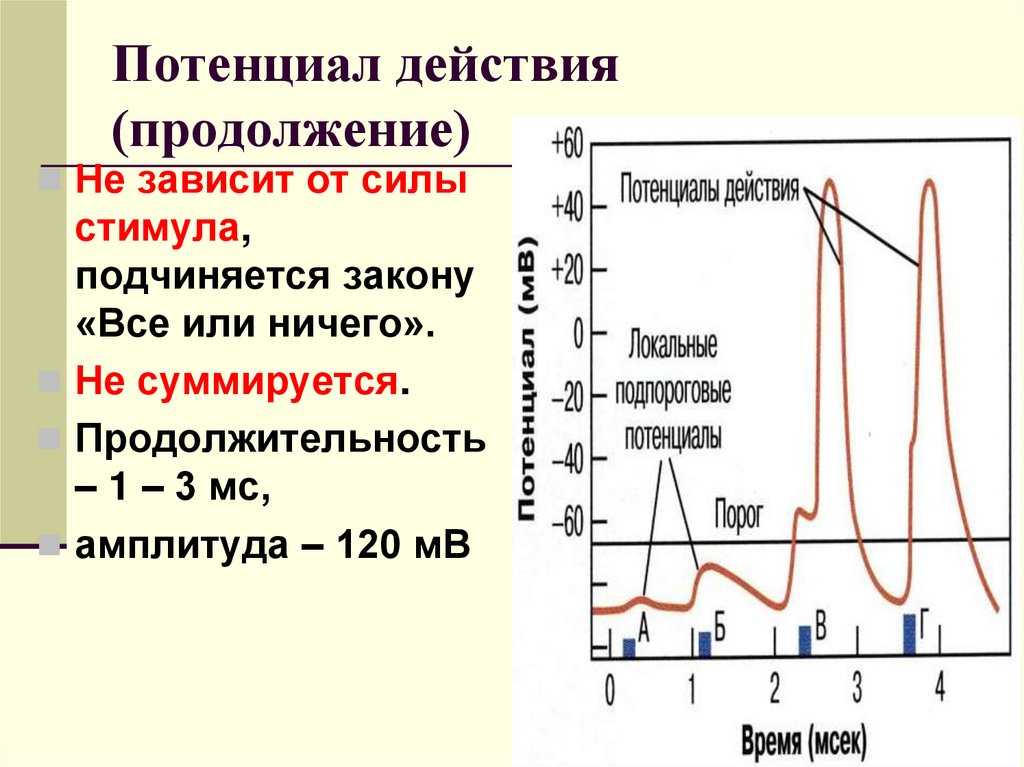
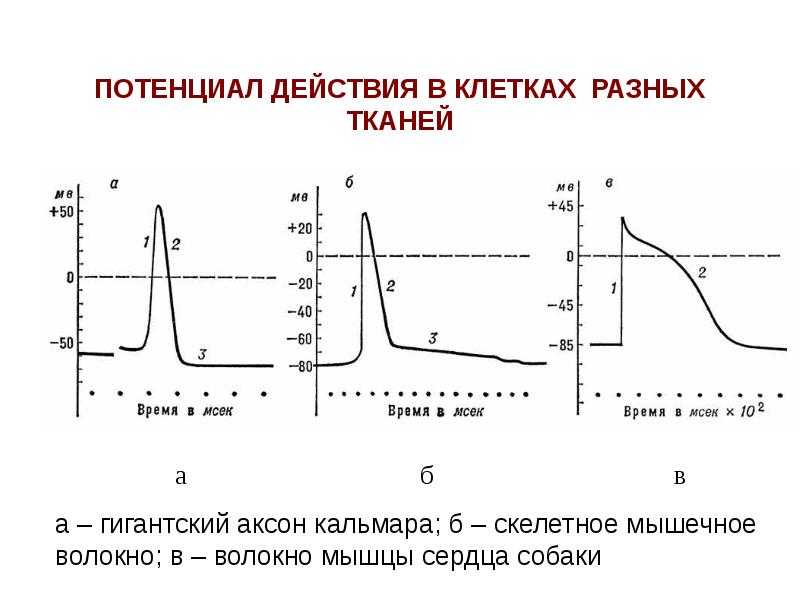
Интересно, что амплитуда потенциала действия у каждой клетки постоянная и своя, правда, колеблется не так сильно и в основном составляет по абсолютной величине около 90-110 мВ. А вот длительность ПД у разных типов клеток различается и довольно значительно, вплоть до нескольких порядков.
Наиболее «быстрострельные» потенциалы действия у крупных миелинизированных аксонов двигательных нейронов (2-3 мс), а также скелетных мышечных волокон (3-4 мс). Оно и понятно – им важна скорость. Прежде всего, для выживания.
![Глава x. физиология нервно-мышечной системы [1981 георгиева с.а., беликина н.в., прокофьева л.и., коршунов г.в., киричук в.ф., головченко в.м., токаева л.к. - физиология человека]](https://mapisa-plitka.ru/wp-content/uploads/e/a/f/eaf48efa22aebe920b8930916ab30e1b.jpeg)

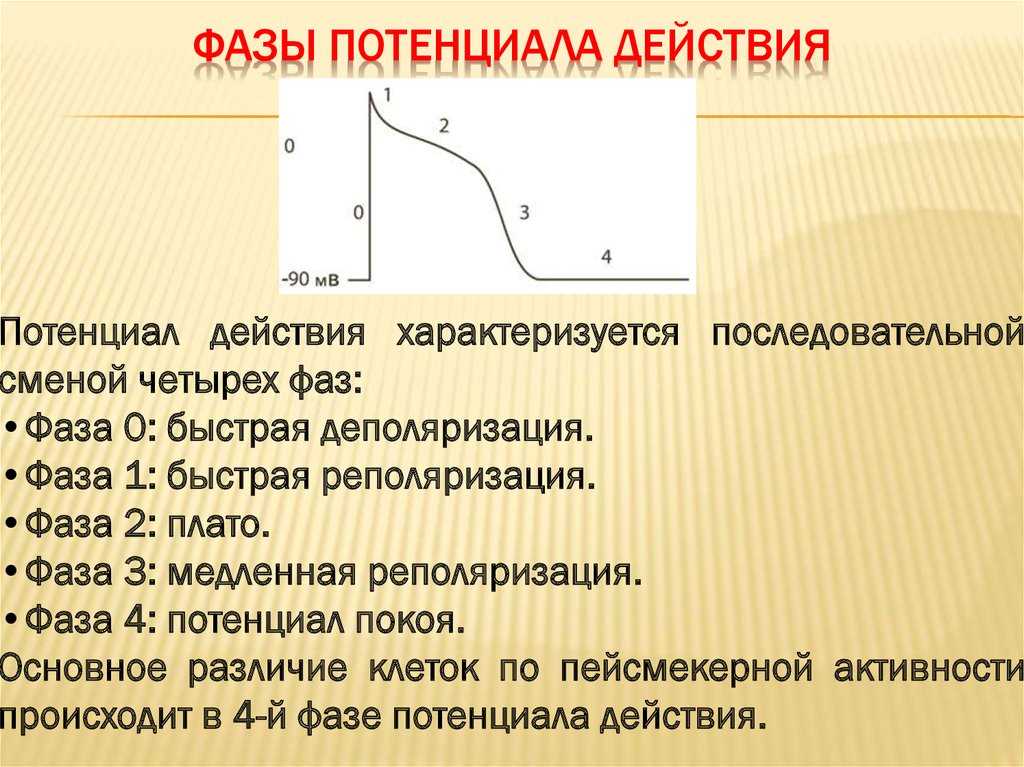
Гладкомышечные клетки генерируют потенциалы действия не всегда, а их продолжительность может составлять от 10 до 30 мс, как, например, в клетках желудочно-кишечного тракта. Здесь сила сокращения мышцы зависит от того, насколько часто возникают серийные потенциалы. Схема такая: чем чаще один ПД следует за другим, тем больше кальция поступает в клетки, и, соответственно, тем сильнее сокращается и тонизируется гладкая мышца.
У кардиомиоцитов (мышечных клеток сердца) длительность ПД больше на целых два порядка и составляет 300-400 мс, так как здесь во время второй фазы потенциала внутрь поступают ионы кальция, которые нужны для того, чтобы запустить сокращение. В итоге чем длительность ПД отдельной клетки выше, тем сильнее сокращение сердца.
Зачем нам всё это нужно знать? Всё достаточно просто. Потенциал действия и колебания потенциала покоя лежат в основе возбуждения и торможения у животных и человека. Главные характеристики ПД – амплитуда и длительность – могут быть как функциональными, так и носить патологический характер, что сразу даст нам понять о «сбоях в системе». А исследование суммарных потенциалов действия клеток и органов легло в основу таких незаменимых в современной врачебной практике диагностических методик, как электрокардиография, электроэнцефалография и электромиография.
Текст: Анна Хоружая
Предыдущие выпуски:
как появляется нервная система,
клетки нервной системы,
Общие положения[]
Файл:Action potential RUS.png
Рис. 2. A. Схематичное изображение идеализированного потенциала действия. B. Реальный потенциал действия пирамидного нейрона гиппокампа крысы. Форма реального потенциала действия обычно отличается от идеализированной.
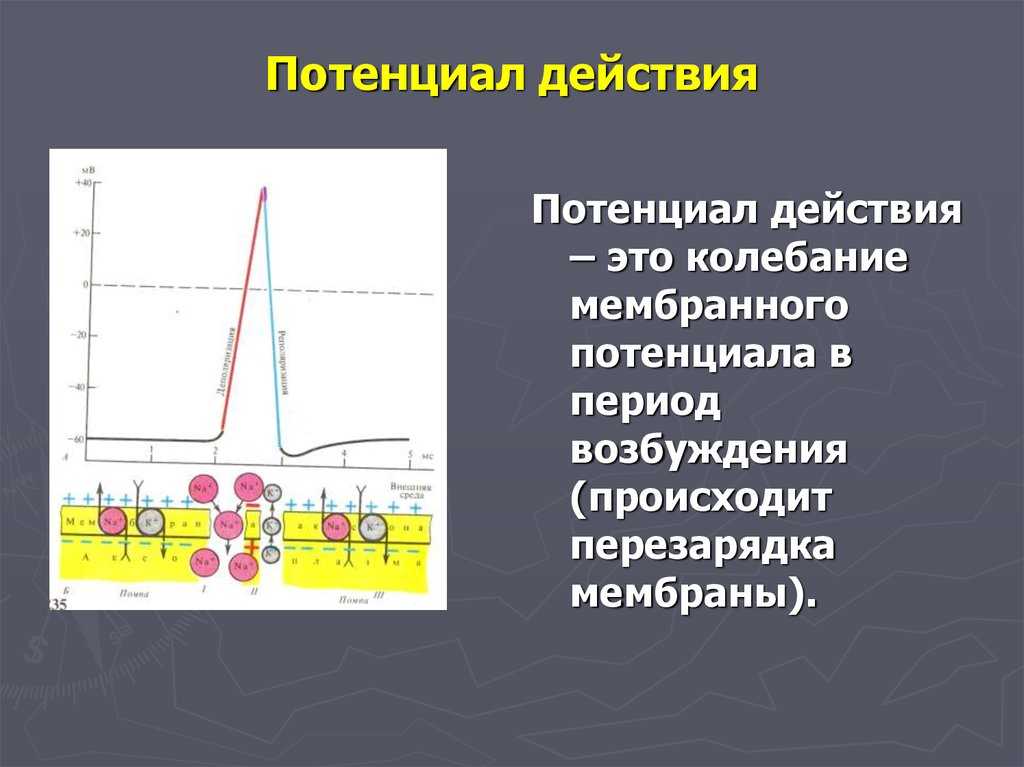
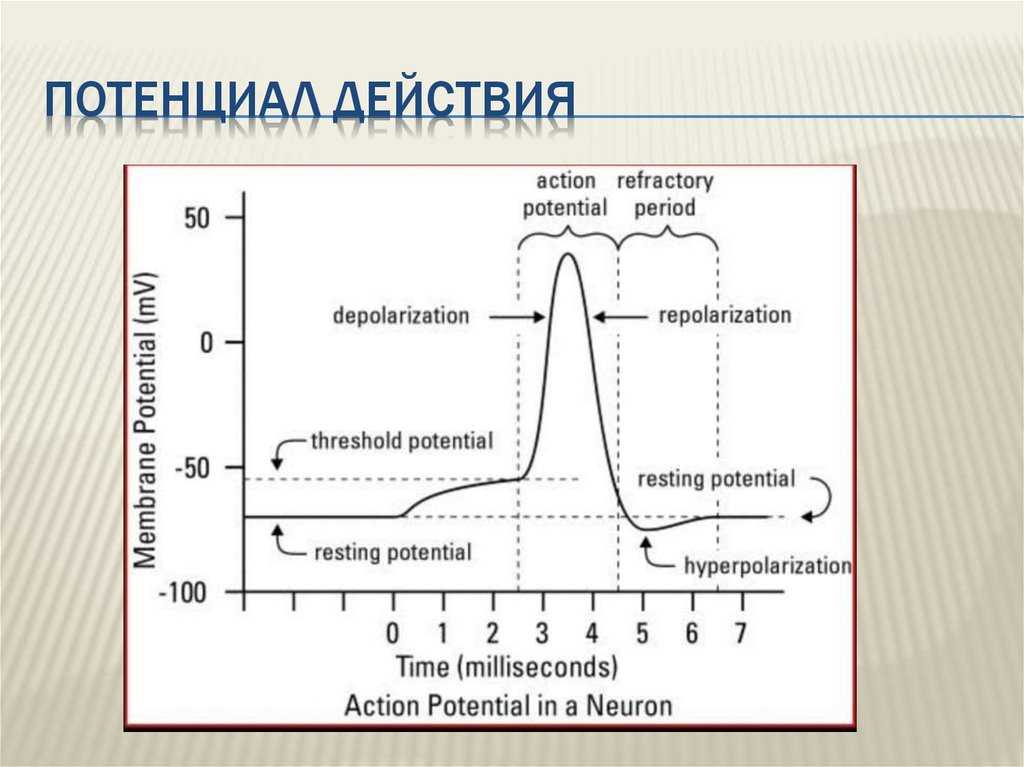
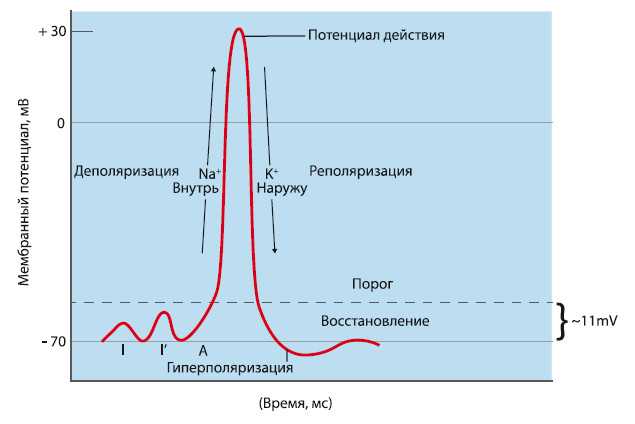
Поляризация мембраны живой клетки обусловлена отличием ионного состава с её внутренней и наружной стороны. Когда клетка находится в спокойном (невозбуждённом) состоянии, ионы по разные стороны мембраны создают относительно стабильную разность потенциалов, называемую потенциалом покоя. Если ввести внутрь живой клетки электрод и измерить мембранный потенциал покоя, он будет иметь отрицательное значение (порядка −70 — −90 мВ). Это объясняется тем, что суммарный заряд на внутренней стороне мембраны существенно меньше, чем на внешней, хотя с обеих сторон содержатся и катионы, и анионы. Снаружи — на порядок больше ионов натрия, кальция и хлора, внутри — ионов калия и отрицательно заряженных белковых молекул.
Потенциал мембраны может изменяться под действием различных стимулов. Искусственным стимулом может служить электрический ток, подаваемый на внешнюю или внутреннюю сторону мембраны через электрод. В естественных условиях стимулом часто служит химический сигнал от соседних клеток, поступающий через синапс или путём диффузной передачи через межклеточную среду. Смещение мембранного потенциала может происходить в отрицательную (гиперполяризация) или положительную (деполяризация) сторону.

![Глава x. физиология нервно-мышечной системы [1981 георгиева с.а., беликина н.в., прокофьева л.и., коршунов г.в., киричук в.ф., головченко в.м., токаева л.к. - физиология человека]](https://mapisa-plitka.ru/wp-content/uploads/b/e/6/be6c0145427c8341b51e2c827705784d.jpeg)
В нервной ткани потенциал действия как правило возникает при деполяризации — если деполяризация мембраны нейрона достигает некоторого порогового уровня или превышает его, клетка возбуждается и от ее тела к аксонам и дендритам распространяется волна электрического сигнала.
Файл:Натриевые каналы.jpg
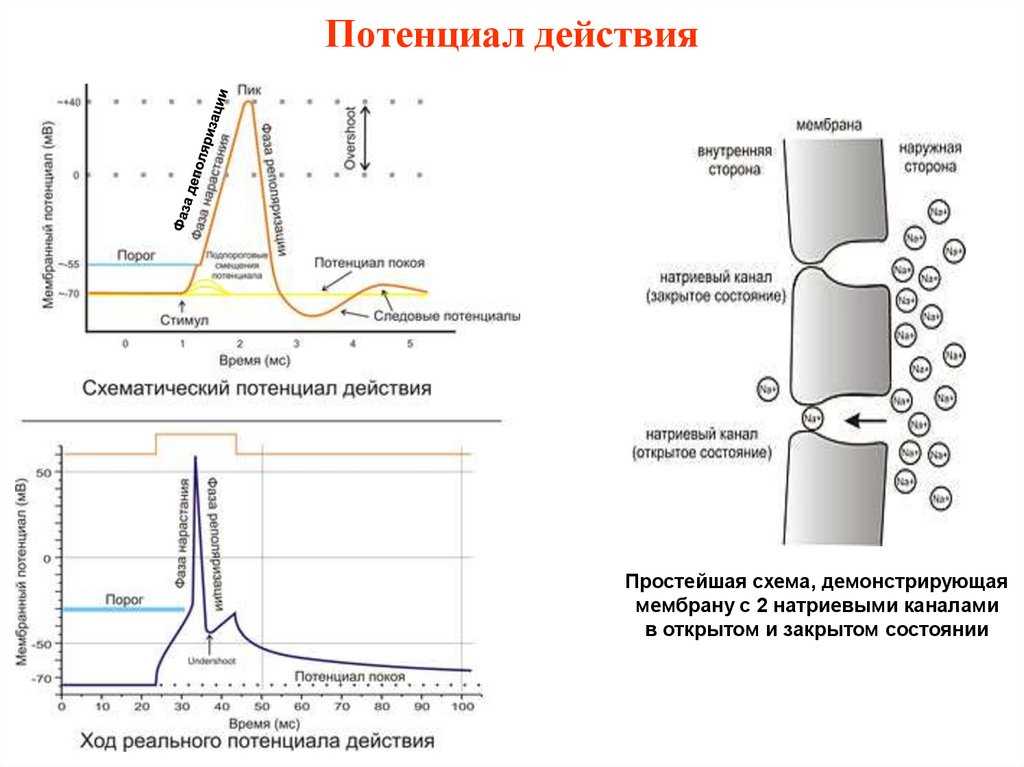
Рис. 3. Простейшая схема, демонстрирующая мембрану с двумя натриевыми каналами в открытом и закрытом состоянии, соответственно
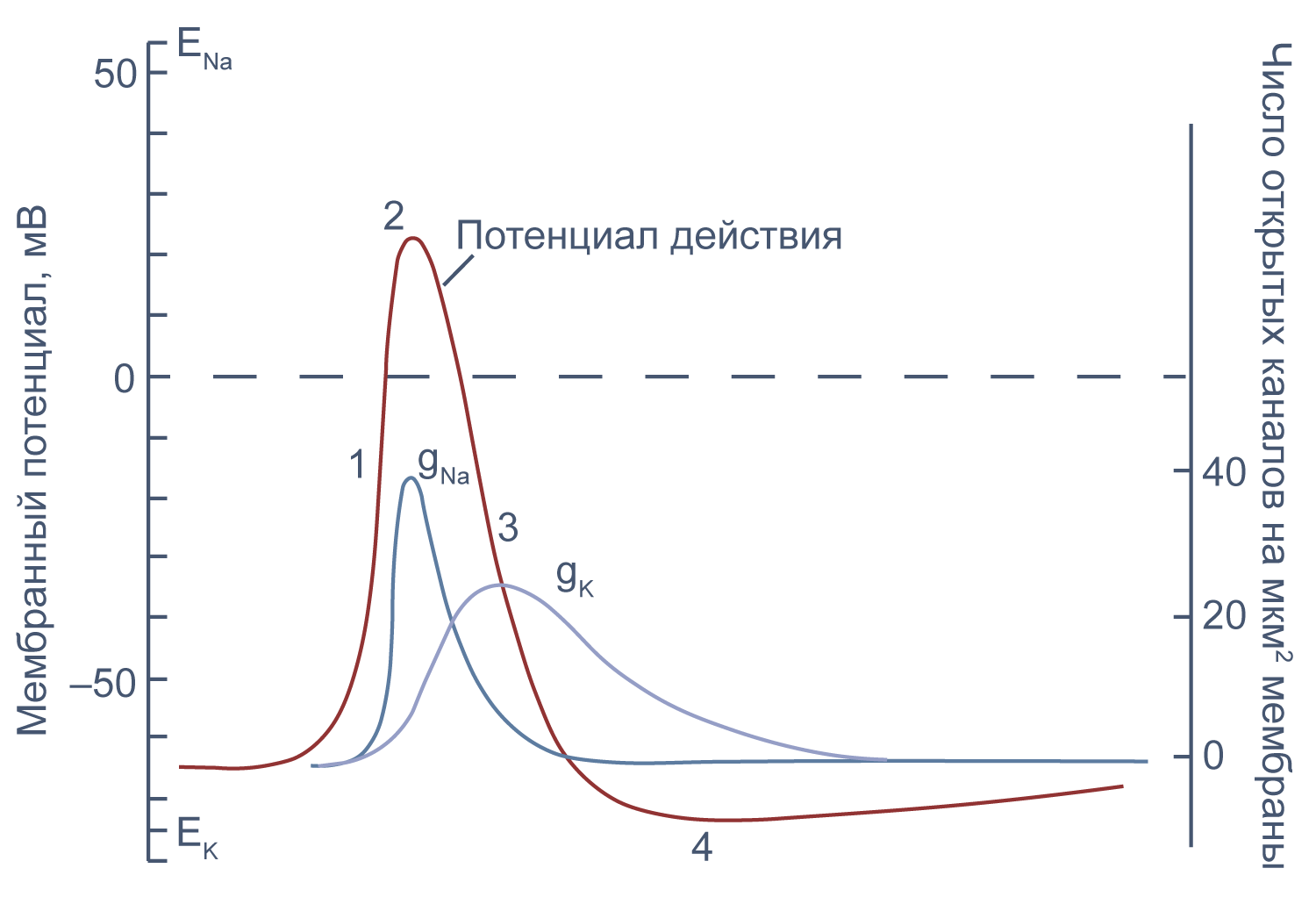
Это обусловлено тем, что на мембране клетки находятся ионные каналы — белковые молекулы, образующие в мембране поры, через которые ионы могут проходить с внутренней стороны мембраны на наружную и обратно. Большинство каналов ионоспецифично — то есть, натриевый канал пропускает практически только ионы натрия, и не пропускает другие (это явление называют селективностью). Мембрана клеток возбудимых тканей (нервной и мышечной) содержит большое количество потенциал-зависимых ионных каналов — способных быстро реагировать на смещение мембранного потенциала. Деполяризация мембраны в первую очередь вызывает открытие потенциал-зависимых натриевых каналов. Когда одновременно открывается достаточно много натриевых каналов, положительно заряженные ионы натрия устремляются через них на внутреннюю сторону мембраны. Движущая сила в данном случае обеспечивается градиентом концентрации (с внешней стороны мембраны находится намного больше положительно заряженных ионов натрия, чем внутри клетки) и отрицательным зарядом внутренней стороны мембраны (см. Рис. 2).
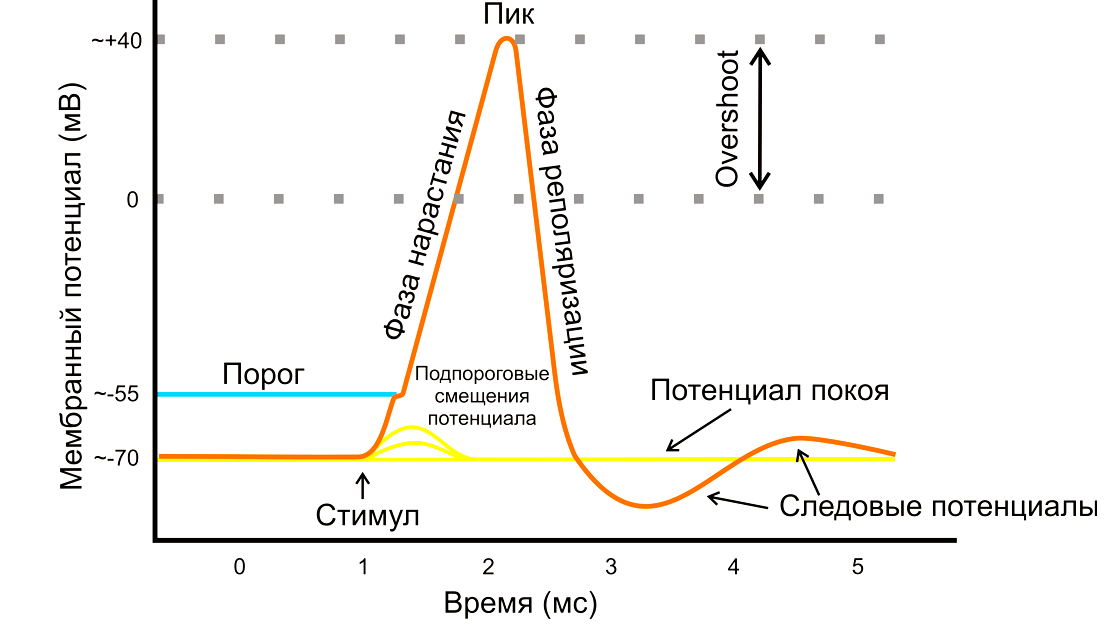
Поток ионов натрия вызывает ещё бо́льшее и очень быстрое изменение мембранного потенциала, которое и называют потенциалом действия (в специальной литературе обозначается ПД).
Согласно закону «всё-или-ничего» мембрана клетки возбудимой ткани либо не отвечает на стимул совсем, либо отвечает с максимально возможной для неё на данный момент силой. То есть, если стимул слишком слаб и порог не достигнут, потенциал действия не возникает совсем; в то же время, пороговый стимул вызовет потенциал действия такой же амплитуды, как и стимул, превышающий пороговый. Это отнюдь не означает, что амплитуда потенциала действия всегда одинакова — один и тот же участок мембраны, находясь в разных состояниях, может генерировать потенциалы действия разной амплитуды.
После возбуждения нейрон на некоторое время оказывается в состоянии абсолютной рефрактерности, когда никакие сигналы не могут его возбудить снова, затем входит в фазу относительной рефрактерности, когда его могут возбудить исключительно сильные сигналы (при этом амплитуда ПД будет ниже, чем обычно). Рефрактерный период происходит из-за инактивации быстрого натриевого тока, то есть инактивации натриевых каналов (см. ниже).
Распространение потенциала действия[]
Распространение потенциала действия по немиелинизированным волокнам
По немиелинизированному волокну ПД распространяется непрерывно. Проведение нервного импульса начинается с распространением электрического поля. Возникший ПД за счет электрического поля способен деполяризовать мембрану соседнего участка до критического уровня, в результате чего на соседнем участке генерируются новые ПД. Сами ПД не перемещаются, они исчезают там же, где возникают. Главную роль в возникновении нового ПД играет предыдущий.
Если внутриклеточным электродом раздражать аксон посередине, то ПД будет распространяться в обоих направлениях. Обычно же ПД распространяется по аксону в одном направлении (от тела нейрона к нервным окончаниям), хотя деполяризация мембраны происходит по обе стороны от участка, где в данный момент возник ПД. Одностороннее проведение ПД обеспечивается свойствами натриевых каналов — после открывания они на некоторое время инактивируются и не могут открыться ни при каких значениях мембранного потенциала (свойство рефрактерности). Поэтому на ближнем к телу клетки участке, где до этого уже «прошел» ПД, он не возникает.
При прочих равных условиях распространение ПД по аксону происходит тем быстрее, чем больше диаметр волокна. По гигантским аксонам кальмара ПД может распространяться почти с такой же скоростью, как и по миелинизированным волокнам позвоночных (около 100 м/c).
Распространение потенциала действия по миелинизированным волокнам
Распространение мембранного потенциала
По миелинизированному волокну ПД распространяется скачкообразно (сальтаторное проведение). Для миелинизированных волокон характерна концентрация потенциалзависимых ионных каналов только в областях перехватов Ранвье; здесь их плотность в 100 раз больше, чем в мембранах безмиелиновых волокон. В области миелиновых муфт потенциалзависимых каналов почти нет. ПД, возникший в одном перехвате Ранвье, за счет электрического поля деполяризует мембрану соседних перехватов до критического уровня, что приводит к возникновению в них новых ПД, то есть возбуждение переходит скачкообразно, от одного перехвата к другому. В случае повреждения одного перехвата Ранвье ПД возбуждает 2-ой, 3-ий, 4-ый и даже 5-ый, поскольку электроизоляция, создаваемая миелиновыми муфтами, уменьшает рассеивание электрического поля. Это увеличивает скорость распространения ПД по миелинизированным волокнам по сравнению с немиелинизированными. Кроме того, миелинизированные волокна толще, а электрическое сопротивление более толстых волокон меньше, что тоже увеличивает скорость проведения импульса по миелинизированным волокнам. Другим преимуществом сальтаторного проведения является его экономичность в энергетическом плане, так как возбуждаются только перехваты Ранвье, площадь которых меньше 1 % мембраны, и, следовательно, необходимо значительно меньше энергии для восстановления трансмембранных градиентов Na+ и K+, расходующихся в результате возникновения ПД, что может иметь значение при высокой частоте разрядов, идущих по нервному волокну.


Чтобы представить, насколько эффективно может быть увеличена скорость проведения за счёт миелиновой оболочки, достаточно сравнить скорость распространения импульса по немиелинизированным и миелинизированным участкам нервной системы человека. При диаметре волокна около 2 µм и отсутствии миелиновой оболочки скорость проведения будет составлять ~1 м/с, а при наличии даже слабой миелинизации при том же диаметре волокна — 15-20 м/с. В волокнах большего диаметра, обладающих толстой миелинововой оболочкой, скорость проведения может достигать 120 м/с.
Следует однако понимать, что скорость распространения потенциала действия по мембране отдельно взятого нервного волокна отнюдь не является постоянной величиной — в зависимости от различных условий, эта скорость может очень значительно уменьшаться и, соответственно, увеличиваться, возвращаясь к некоему исходному уровню.